
- Title
-
Essential role of lysyl oxidases in notochord development
- Authors
- Gansner, J.M., Mendelsohn, B.A., Hultman, K.A., Johnson, S.L., and Gitlin, J.D.
- Source
- Full text @ Dev. Biol.
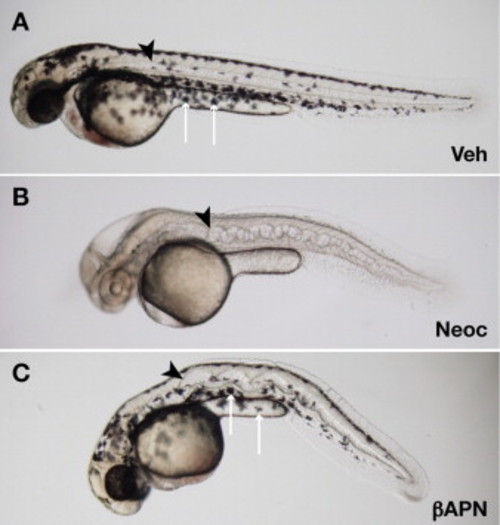
β-Aminopropionitrile recapitulates the notochord phenotype of copper deficiency. Wild-type embryos were incubated in vehicle (A), 10 μM neocuproine (B), or 10 mM β-aminopropionitrile supplemented with 10 �M CuCl2 (C). At 48 hpf, the notochord (arrowheads) is distorted in embryos treated with neocuproine (B) and β-aminopropionitrile (C); however, melanocytes (arrows) are present after β-aminopropionitrile (C) but not neocuproine treatment (B), demonstrating that β-aminopropionitrile does not act through copper chelation. |
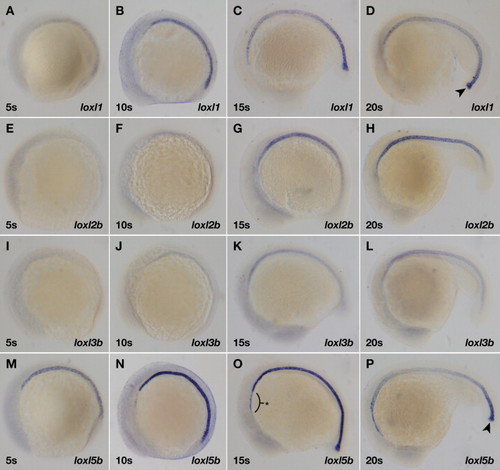
Four lysyl oxidases are expressed throughout the developing zebrafish notochord by in situ hybridization. (A?D) loxl1; (E?H) loxl2b; (I?L) loxl3b; and (M?P) loxl5b. Stages of development are 5 somites (A, E, I, M); 10 somites (B, F, J, N); 15 somites (C, G, K, O); and 20 somites (D, H, L, P). loxl1 and loxl5b are robustly expressed at the caudal notochord tip (arrowheads) while loxl5b is also expressed anterior to the notochord (asterisk). EXPRESSION / LABELING:
|
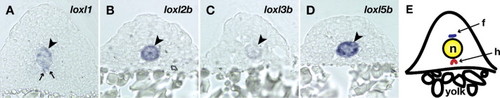
Lysyl oxidases are expressed by notochord vacuolar cells. (A?D) Frozen cross-sections of 15 somite embryos were obtained after in situ hybridization with the following probes: (A) loxl1; (B) loxl2b; (C) loxl3b; and (D) loxl5b. The notochord is indicated (arrowheads). Hypochord staining of loxl1 (arrows) suggests a role for this family member in extracellular matrix crosslinking near the developing aorta. (E) Cartoon demonstrating the location of the hypochord (h) and floorplate (f), which are closely apposed to the notochord (n). Lysyl oxidases are not expressed in the floorplate. EXPRESSION / LABELING:
|
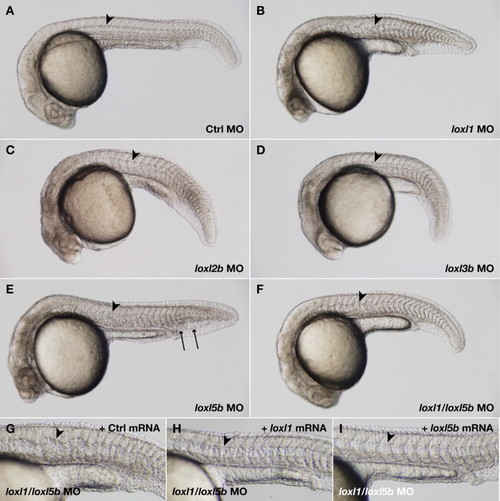
Morpholino knockdown of loxl1 or loxl1 and loxl5b together results in notochord distortion. Wild-type embryos were injected with morpholinos (MO) to notochord-expressed lysyl oxidases and photographed at 24 hpf. (A) 12 ng standard control; (B) 6 ng loxl1; (C) 3.7 ng loxl2b; (D) 12 ng loxl3b; (E) 7.4 ng loxl5b; (F) 2.4 ng loxl1 and 5 ng loxl5b. The notochord (arrowheads) is strikingly distorted after knockdown of loxl1 (B), and caudal vein edema develops with the loxl5b morpholino (E, arrows). Combining doses of loxl1 and loxl5b morpholino that do not cause notochord distortion alone also recapitulates the notochord phenotype seen with neocuproine and β-aminopropionitrile treatment (F). This distortion is specifically rescued by co-injection of mRNA encoding either loxl1 (H) or loxl5b (I), but not control sequence (G). PHENOTYPE:
|
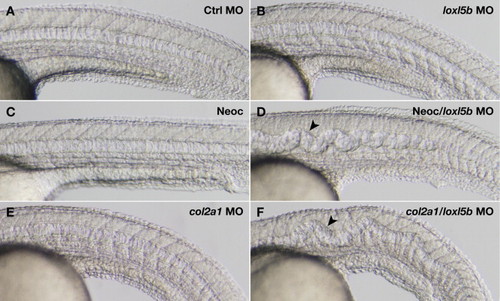
Partial knockdown of loxl5b sensitizes embryos to notochord distortion in the presence of suboptimal copper nutrition (A?D) or disruption of col2a1 (E, F). Wild-type embryos were injected with 5 ng of control morpholino (A, C) or loxl5b morpholino (B, D), and incubated with (C, D) or without (A, B) 2 μM neocuproine starting at 3 hpf. Embryos injected with loxl5b morpholino develop notochord distortion (arrowhead) in the context of diminished copper availability (D). Embryos injected with 7.4 ng of col2a1 morpholino (E) are sensitized to develop notochord distortion (arrowhead) upon co-injection of 5 ng of loxl5b morpholino (F), demonstrating a genetic interaction between col2a1 and loxl5b. Photographs were obtained at 30 hpf (A?D) and 24 hpf (E, F). |
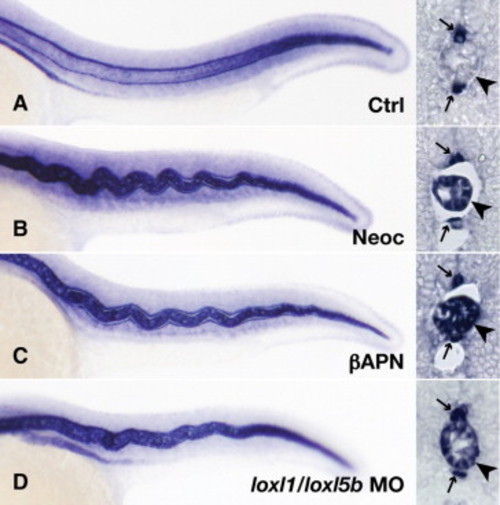
Expression of col2a1 mRNA by notochord vacuolar cells persists late following lysyl oxidase inhibition. Wild-type embryos were injected with 7.4 ng control morpholino (A), incubated in 10 μM neocuproine (B), incubated in 10 mM β-aminopropionitrile supplemented with 10 μM CuCl2 (C), or injected with 3.7 ng each of loxl1 and loxl5b morpholino (D). In situ hybridization was carried out at 24 hpf, and frozen sections confirmed persistent col2a1 expression following lysyl oxidase inhibition (B?D, arrowhead), as well as floorplate and hypochord staining (A?D, arrows). |
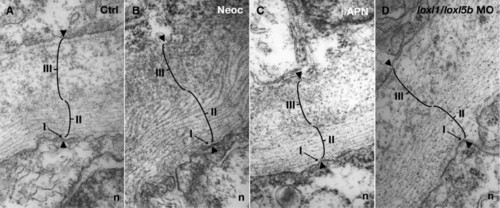
Electron micrographs of the notochord sheath from cross-sections of 30 hpf embryos: (A) 7.4 ng control morpholino; (B) 10 μM neocuproine; (C) 10 mM β-aminopropionitrile; (D) 3.7 ng each of loxl1 and loxl5b morpholino. Sheath components include a basal lamina (I), fibrillar layer (II), and granular layer (III). All three layers are preserved after lysyl oxidase inhibition. |
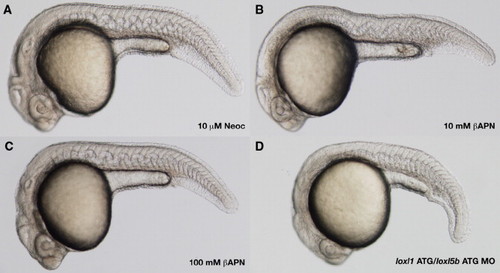
Comparison of the notochord distortion obtained in zebrafish using three distinct methods of lysyl oxidase inhibition at 24 hpf. Wild-type embryos were incubated in 10 μM neocuproine (A), 10 mM �-aminopropionitrile supplemented with 10 μM CuCl2 (B), 100 mM �-aminopropionitrile supplemented with 10 μM CuCl2 (C), or injected with 6 ng each of loxl1 and loxl5b start (ATG) morpholino (D). 10 μM neocuproine (A) causes greater notochord distortion than 10 mM �-aminopropionitrile (B), but an identical phenotype is obtained with 100 mM �-aminopropionitrile (A vs. C). Melanin pigmentation has not yet developed in the embryos pictured, but is not affected by �-aminopropionitrile treatment or by the morpholino injection. PHENOTYPE:
|
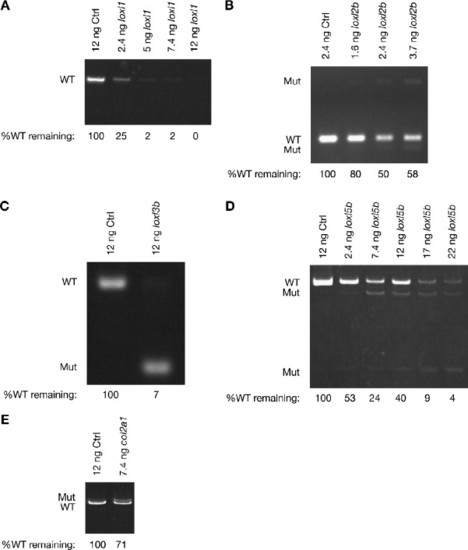
Lysyl oxidase and collagen II splice morpholinos decrease the abundance of wild-type splice forms and cause specific changes in splice site usage. (A) loxl1, (B) loxl2b, (C) loxl3b, (D) loxl5b and (E) col2a1. Wild-type embryos injected with the indicated doses of morpholino were harvested at 20 hpf and cDNA prepared from equal amounts of RNA. The intensity of the wild-type (WT) product obtained by PCR was quantified and the percentage of wild-type splice form remaining after morpholino knockdown determined, as noted. This measure is independent of processes that may depress levels of the mutant (Mut) splice products, including degradation through nonsense-mediated decay. The primer set used to amplify loxl1 does not amplify the mutant band. EXPRESSION / LABELING:
|
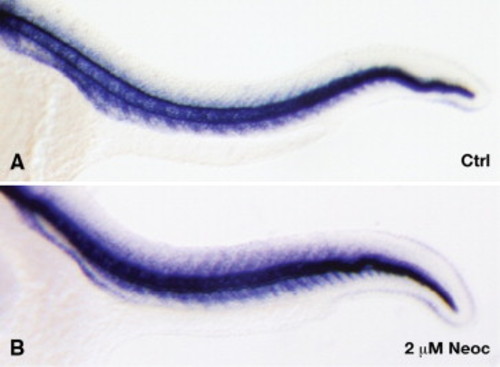
Persistent expression of col2a1 by notochord vacuolar cells after lysyl oxidase inhibition is not due to mechanical distortion of the notochord. Wild-type embryos incubated with (B) or without (A) 2 μM neocuproine were probed for col2a1 expression at 24 hpf. col2a1 expression persists in the notochord vacuolar cells after treatment with 2 μM neocuproine, which affects crosslinking but does not cause mechanical distortion. This suggests that the signal to downregulate col2a1 expression in the vacuolar cells of the notochord originates from the products of the crosslinking reaction. EXPRESSION / LABELING:
|

Unillustrated author statements EXPRESSION / LABELING:
|
Reprinted from Developmental Biology, 307(2), Gansner, J.M., Mendelsohn, B.A., Hultman, K.A., Johnson, S.L., and Gitlin, J.D., Essential role of lysyl oxidases in notochord development, 202-213, Copyright (2007) with permission from Elsevier. Full text @ Dev. Biol.