
- Title
-
Common genetic control of haemangioblast and cardiac development in zebrafish
- Authors
- Peterkin, T., Gibson, A., and Patient, R.
- Source
- Full text @ Development

gata4, gata5 and gata6 are essential for anterior haemangioblast specification. (A) gata4, gata5 and gata6 are expressed in overlapping yet distinct domains throughout the anterior lateral plate mesoderm (ALM). (B) Expression of l-plastin, mpx and ikaros, genes associated with myelopoiesis, was absent in gata5+gata6 morphants at 24 hpf. (C) Analysis of earlier regulatory myeloid gene expression on knock down of gata5 and gata6 shows ablation of cmyb, ikaros and pu1 at 10 somites (red arrowheads). (B,C) The number of embryos exhibiting this phenotype shown at the bottom right-hand corner of each panel. (D) Expression of haemangioblast-associated genes close to the onset of their expression was downregulated in gata5+gata6 morphants (marked by red arrowheads where necessary). uncx4.1 was used as a somite marker to stage the embryos (white brackets). n=40-100 for each gene analysed, the pictures shown depict >95% of the embryos. Views are: (A) flatmounted, anterior to the top; (B) lateral, anterior to left, except for ikaros, which is an anterior view; (C) dorsal, anterior to the top; (D) anterior-dorsal. EXPRESSION / LABELING:
PHENOTYPE:
|

gata5 and gata6 rescue and gain-of-function experiments confirm necessity and identify sufficiency for myelopoiesis. Injection of exogenous gata5 and gata6 mRNA (g5+6RNA) into wild-type embryos did not induce fli1 or etsrp, whereas scl and pu1 were ectopically expressed. Rescue of all four genes was seen in gata5 and gata6 morphants co-injected with gata5 and gata6 mRNA (g5+6mo + g5+6RNA) when compared with gata5 and gata6 morphants alone (g5+6mo). Views are anterior-dorsal. EXPRESSION / LABELING:
|
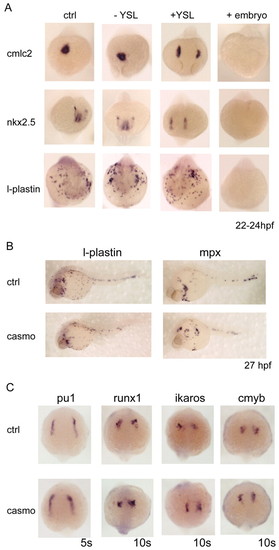
Depletion of gata5 and gata6 from the YSL causes cardia bifida but does not affect myelopoiesis. (A) The YSL expression of gata5 and gata6 was depleted by injection of gata5+gata6 morpholinos into the YSL at the 1000-cell stage. To trace the correctly targeted embryos, a fluorescent control morpholino was co-injected. Embryos positive for fluorescence in the YSL alone (+YSL) were selected at several time points and harvested as YSL gata5+gata6-depleted embryos. The embryos showing no fluorescence (-YSL) were collected as negative injection controls and should express wild-type levels of gata5 and gata6. To ensure the efficiency of the morpholinos, positive control embryos were injected with gata5+gata6 morpholinos at the one-cell stage and were fluorescent throughout the embryo (+embryos). To assess heart formation, cmlc2 and nkx2.5 expression was analysed. Depletion of gata5 and gata6 in the YSL (+YSL) showed normal levels of expression of both cmlc2 and nkx2.5, indicating that specification occurs normally in these embryos. However, cardia bifida was observed in the +YSL embryos, demonstrating that gata5 and gata6 are required in the YSL for the correct migration of the cardiac precursors to the midline. By contrast, l-plastin expression was the same as in the wild-type embryos, indicating that gata5 and gata6 expression in the YSL is dispensable for myelopoiesis. Embryos injected at the one-cell stage (+embryos) showed complete absence of cmlc2, nkx2.5 and l-plastin expression, thereby validating morpholino effectiveness. Views are anterior. For each gene and type (�YSL, +embryo) analysed, n=28-38, and the images shown depict the findings for >95% of the embryos. (B,C) Loss of endoderm in casanova morphant embryos but no defects in myelopoiesis. To establish whether endoderm plays a role in myelopoiesis, endodermless embryos were created by injection of casanova morpholinos. Casanova morphants (casmo) were assessed for endoderm formation and myelopoiesis. Myelopoiesis was not affected in cas morphants as l-plastin and mpx remained unaffected (B). Gene expression in the ALM of cas morphants was examined at 5 and 10 somites (C). The formation of myeloid precursors occurred as normal in cas morphants. Expression of pu1 at 5 somites, and runx1, ikaros and cmyb at 10 somites, was unaffected in cas morphants. Views are anterior-dorsal. For each gene analysed n=38-67, and the images shown depict the findings for >95% of the embryos. EXPRESSION / LABELING:
PHENOTYPE:
|

Initiation of gata4, gata5 and gata6 expression is independent of scl and cloche. (A) Expression of the GATA factors was analysed in scl morphants. gata4, gata5 and gata6 were expressed as in the wild-type embryos in the ALM (red arrowheads), placing them upstream of scl. Loss of pu1 was used as a control for scl morpholino efficacy. (B) The expression of gata4, gata5 and gata6 in the ALM was unchanged (red arrowheads) in cloche embryos placing the GATA factors parallel to or upstream of cloche. (C) cloche embryos were identified by the absence of gata1 expression (red arrowheads) in the PLM. Pu1 downregulation was an additional control (B). The expression of uncx4.1 was used as a somitic counter to ensure correct staging of the embryos (white brackets). Views are anterior-dorsal. For each gene analysed, n=28-42, and the images shown depict the findings for >95% of the embryos identified as cloche and scl morphant embryos. EXPRESSION / LABELING:
|
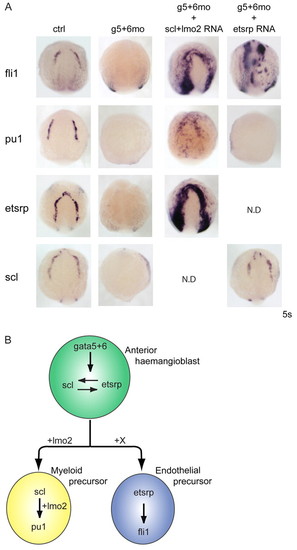
Interaction of gata5 and gata6 with scl/lmo2 and etsrp. (A) Scl/lmo2 mRNA can rescue both blood (pu1), and endothelium (fli1 and etsrp) development in gata5 and gata6 morphants at the 5 somite stage. By contrast, etsrp overexpression can rescue fli1 but not pu1 expression in gata5 and gata6 morphants. All views are anterior. N.D., not determined. (B) Proposed model for gata5 and gata6 function within the anterior haemangioblast. gata5 and gata6 are required for etsrp and scl expression in the haemangioblast. gata5 and gata6 overexpression can induce scl and pu1 expression, which drives the myeloid lineage, whereas endothelium cannot be induced by gata5 and gata6 mRNA (see Fig. 2). Furthermore, the head vasculature eventually recovers (see Fig. S4A in the supplementary material), suggesting the existence of an unknown GATA-independent signal (X). EXPRESSION / LABELING:
|
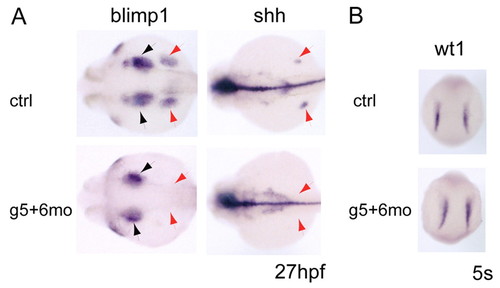
Fate of ALM cells in gata5 and gata6 morphants. (A) Fin buds are lost in gata5 and gata6 morphants, as seen by the loss of blimp1 and shh expression (red arrows). Pharyngeal endoderm is slightly reduced in morphants (blimp1, black arrows) compared with wild type. (B) Pronephric mesoderm (wt1 expression) remains unchanged in gata5 and gata6 morphants at 5 somites. EXPRESSION / LABELING:
PHENOTYPE:
|
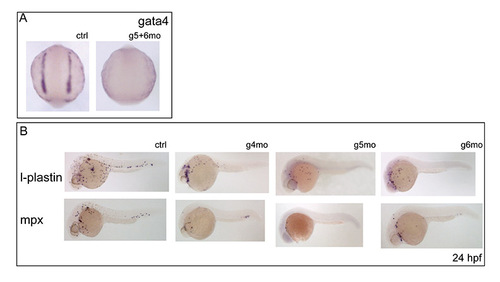
Defects in the ALM and its derivatives on loss of gata4, gata5 and/or gata6. (A) gata4 is downregulated in gata5+gata6 morphants at 5 somites, showing that in effect gata5+gata6 morphants are a triple knockdown model. (B) Myelopoiesis is reduced but not abolished in single GATA morphants as seen by a downregulation of l-plastin and mpx in gata4, gata5 and gata6 morphants. |
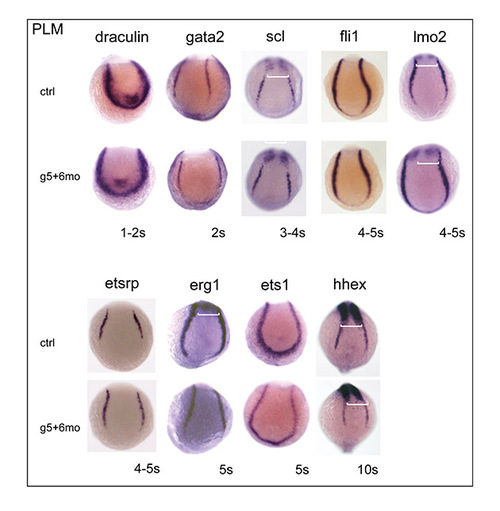
PLM expression of haemangioblast-associated genes is independent of gata4, gata5 and gata6. The expression of haemangioblast-associated genes was analysed close to the onset of their expression. Expression in the PLM was unchanged in gata5+gata6 morphants. uncx4.1 was used as a somite marker to stage the embryos (white brackets) and the stage of the embryos is shown under each panel. Posterior views. For each gene analysed, n=40-100, the images shown represent the findings for >95% of the embryos. |
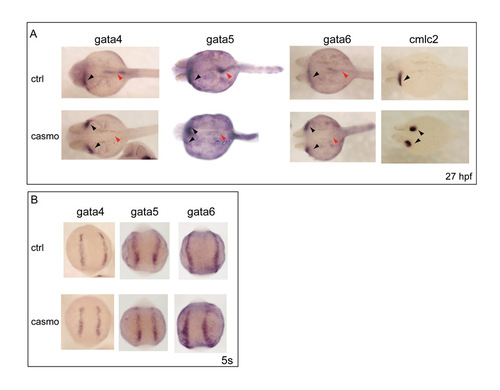
gata4, gata5 and gata6 expression is lost in the endoderm but not in the mesoderm of casanova morphants. (A) Expression of gata4, gata5 and gata6 was downregulated in the gut tube of cas morphants (red arrows) at 27 hpf, whilst expression remained in the cardiac regions (black arrowheads). Cardia bidifa was observed in cas morphants (two black arrowheads), whilst wild-type embryos possess a single heart tube (one black arrowhead). Cardia bifida in cas morphants was confirmed by the bilateral expression of cmlc2, which is not seen in control embryos. (B) Expression of gata4, gata5 and gata6 at 5 somites remained normal in the ALM of casanova morphants. Images shown in A are dorsal views, anterior to left; those in B are anterior-dorsal views. |

Pu1 is required for myeloid differentiation but not for anterior haemangioblast formation. (A) Recovery of the head endothelial programme is seen in gata5 and gata6 morphants. fli1 expression appears normal at 22 and 30 hpf. (B,C) To assess the requirement for pu1 in the ALM, embryos were injected with pu1 morpholino (mo) and gene expression was analysed at 5 and 10 somites. (B) The loss of pu1 expression itself in the ALM, and the PLM (data not shown), confirmed pu1 mo efficiency (red arrows). gata2, gata4, gata5 and gata6 were all expressed as in the wild-type embryos in the ALM, as was draculin. Ectopic expression of gata1 was not observed at 5 somites in pu1 morphants. (C) Expression of the haemangioblast-associated genes scl, fli1, lmo2 and hhex was unchanged at 10 somites in pu1 morphants, demonstrating that pu1 is not required for anterior haemangioblast formation. However, cmyb, runx1 and ikaros were specifically lost in the ALM of embryos depleted of pu1 (red arrowheads), confirming a requirement for pu1 in establishing the myeloid programme. Anterior-dorsal views. For each gene analysed, n=43-97, the images shown represent the findings for >95% of the embryos. |