
- Title
-
Tuba1a gene expression is regulated by KLF6/7 and is necessary for CNS development and regeneration in zebrafish
- Authors
- Veldman, M.B., Bemben, M.A., and Goldman, D.
- Source
- Full text @ Mol. Cell Neurosci.
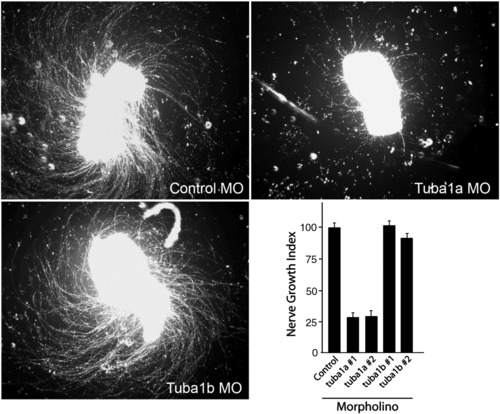
Knockdown of Tuba1a, but not Tuba1b, inhibits RGC axon regeneration in retinal explants. Control, tuba1a or tuba1b-targeting morpholino-modified antisense oligonucleotides (MO) were delivered to RGCs in vivo by placing MO-soaked Gelfoam on the sectioned optic nerve stump. The next day the Gelfoam was removed and 3 days later retinas were isolated, diced and placed in explant culture for 4 days prior to analysis of neurite outgrowth. Shown are representative images of axon outgrowth. The lower right-hand graph shows Nerve Growth Index (see Experimental methods) for control MO and 2 different MOs, #1 and #2, targeting different tuba1a and tuba1b sequences, respectively. Nerve Growth Index values were normalized to control MO-treated samples that were set at 100%. Error bars are standard error of the mean. Note MOs targeting tuba1a, but not tuba1b, dramatically suppress RGC axon regeneration. Experiments were repeated at least 3 times with each MO. |
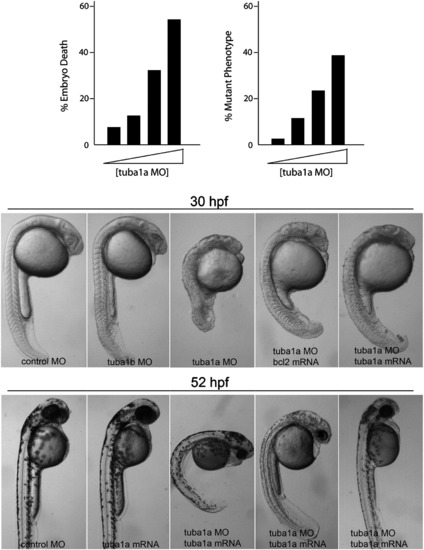
Knockdown of Tuba1a, but not Tuba1b, suppresses CNS formation in developing embryos. Control, tuba1a or tuba1b-targeting morpholinos (MO) were injected into single cell zebrafish embryos. Top graphs show that tuba1a MO-dependent phenotypes at 30 h post fertilization (hpf) are concentration-dependent (tuba1a MO concentrations are 0, 0.065, 0.125 and 0.25 mM; control MO was used to keep total MO concentrations at 0.25 mM). Bottom images show representative pictures of MOinjected (0.125 mM) embryo phenotypes at 30 and 52 hpf. To inhibit apoptosis and examine MO specificity, some experiments included in vitro transcribed bcl2 mRNA (25 pg) or tuba1a mRNA (12.5 pg) that was engineered to escape MO-mediated knockdown. Note that tuba1a, but not tuba1b MO severely disrupted development, especially of anterior structures; however, inclusion of bcl2 or tuba1a mRNA diminished the severity of Tuba1a knockdown. Three different tuba1a mRNA rescue phenotypes are shown at 52 hpf. Similar results were obtained with a second MO targeting different tuba1a and tuba1b sequences. |
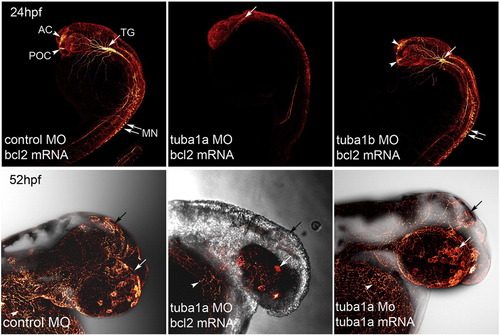
Tuba1a knockdown dramatically inhibits CNS development. Confocal images of zebrafish injected with tuba1a MO and rescue mRNAs. Top panels: Control (0.25 mM), tuba1a (0.125 mM) and tuba1b-targeting morpholinos (MO) (0.25 mM), along with bcl2 mRNA (25 pg) were injected into single cell zebrafish embryos. Embryos were analyzed at 24 h post fertilization for differentiating neurons by whole mount immunostaining with anti-acetylated tubulin antibody. Shown are representative images of immunostained embryos. Arrowheads point to the developing post-optic commissure (POC) and the anterior commissure (AC), while arrows point to the trigeminal ganglion (TG) and the motor neurons (MN). Note the lack of differentiating neurons in the Tuba1a knockdown embryos. Bottom panels: Control (0.25 mM) or tuba1a-targeting (0.125 mM) MOs with bcl2 mRNA (25 pg) or tuba1a mRNA (12.5 pg) were injected into single cell zebrafish embryos. Embryos were analyzed for differentiating neurons by whole mount immunostaining with anti-acetylated tubulin antibody. Shown are representative images of immunostained embryos focusing on the head/eye region. The majority of immunostaining detects sensory neurons on the head (black arrows), eye (white arrows) and running over the yolk (white arrowheads). Note that both the bcl2 and tuba1a mRNA rescued embryos show significant neural differentiation; however, tuba1a mRNA appears to rescue nervous system development much better than bcl2 mRNA. |
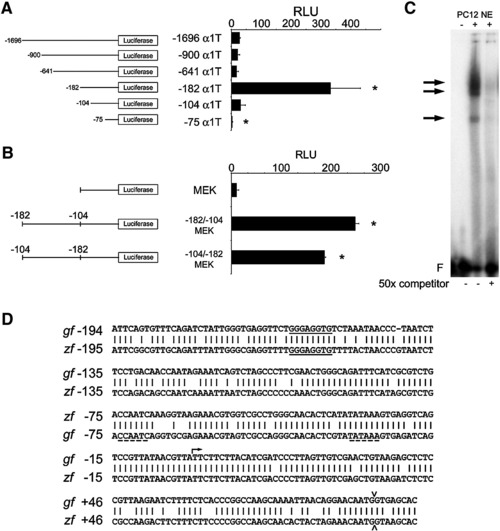
Identification of a tuba1a promoter enhancer. (A) 5′ promoter deletions identify tuba1a regulatory elements. PC12 cells were co-transfected with the indicated tuba1a (α1T)-luciferase constructs along with CMV-CAT for normalization. Two days post-transfection cells were assayed for luciferase and CAT activity. Reported are relative light units (RLU) representing luciferase activity normalized to CAT activity. (*p < 0.05, t-test). (B) The - 182/-104 tuba1a sequence confers activation to a heterologous minimal enkephalin (MEK) promoter in an orientation independent manner. PC12 cells were transfected and assayed, as described in (A), with the indicated constructs. (*p < 0.01, t-test). (C) The - 182/-104 tuba1a enhancer binds proteins present in PC12 nuclear extracts. Radiolabelled - 182/-104 tuba1a enhancer was mixed with and without PC12 nuclear extract (NE) and DNA:protein complexes resolved on native acrylamide gels (EMSA). Shown is a representative autoradiogram indicating free probe (F) and 3 DNA:protein complexes (arrows). Specificity of DNA:protein binding is indicated by competition with 50x unlabeled tuba1a enhancer DNA (competitor). (D) Sequence comparison of the proximal promoter of tuba1a from goldfish (gf) and zebrafish (zf). The conserved protein binding site identified in Fig. 6 is underlined. The CAAT and TATA boxes are identified by dashed underline. An arrow identifies the transcription start site, identified in goldfish, and carrots above and below the sequence identifies the end of exon 1 and beginning of intron 1. Values in (A) and (B) are means +/- SEM. |
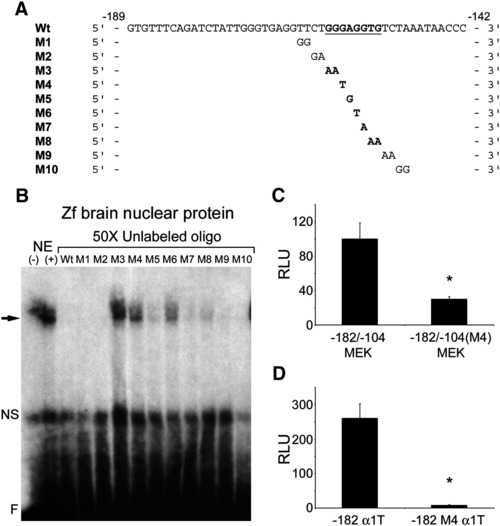
Mutation analysis of the - 182/-104 tuba1a enhancer identifies a G/C-rich sequence that is necessary for nuclear protein binding and enhancer activity. (A) DNA sequences of the oligonucleotides used for mapping the protein binding site in (B). Wt, is the wildtype sequence with the G/C-rich sequence found to be necessary for protein binding typed in bold and underlined. M1?M10, show the basepair changes made in the Wt oligonucleotides for mapping the protein binding enhancer element. (B) EMSA demonstrates that a G/C-rich sequence is necessary for binding nuclear proteins from zebrafish brains. Nuclear protein extract was mixed with radiolabeled Wt oligonucleotides with and without 50x excess of unlabelled mutant oligonucleotides, M1?M10. Specific binding is indicated by a black arrow, nonspecific binding (NS), and free probe (F). Note mutant oligonucleotide competitors M3?M8 partially or completely failed to compete for binding, indicating that these nucleotides are critical for protein binding to the - 182/-104 tuba1a enhancer. (C) A single basepair mutation in the tuba1a enhancer′s G/C-rich sequence reduces enhancer activity in PC12 cells. PC12 cells were transfected with - 182/-104MEK (see Fig. 5B) or - 182/-104(M4)MEK (harbors the - 182/-104 G/C-rich enhancer with the single basepair mutation present in the M4 oligonucleotide, shown in (A), in front of the MEK promoter) along with CMV-CAT for normalization. (D) A single basepair mutation in the G/C-rich protein binding site of the - 182 tuba1a reporter (- 182 α1T in Fig. 5A) reduces promoter activity in PC12 cells. PC12 cells were transfected with - 182 α1T or - 182 M4 α1T (harbors the - 182 α1T sequence with the M4 mutation present in the G/C-rich protein binding site). (C and D) Two days post-transfection cells were assayed for luciferase and CAT activities. Luciferase activity was normalized to CAT activity and reported as relative light units (RLU). Note a single nucleotide substitution in the α1T (tuba1a) promoter′s G/C-rich element decreases enhancer activity. Values are means +/- SEM (*p < 0.01, t-test). |
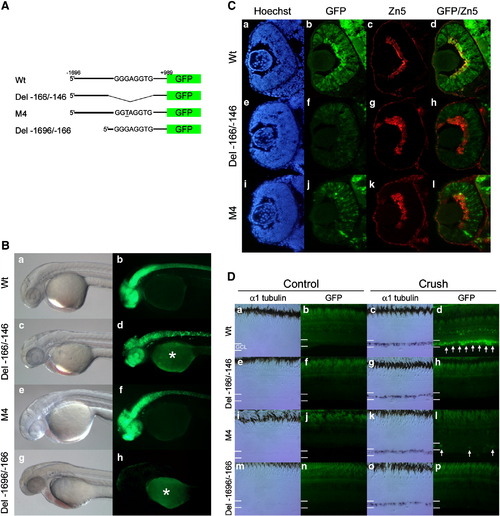
The G/C-rich enhancer is necessary for tuba1a promoter-transgene expression during RGC development and optic nerve regeneration. (A) Schematic representations (not to scale) of the reporter constructs used to create the transgenic lines. Wt transgene includes the 1.696 kb tuba1a promoter with an intact G/C-rich enhancer. Del - 166/-146 transgene contains an internal 20 basepair deletion removing the G/C-rich enhancer. M4 transgene contains the M4 point mutation (underlined) within the G/C-rich enhancer. Del - 1696/-166 transgene contains a deletion of most of the promoter sequence upstream of the G/C-rich enhancer. (B) Representative bright field (a, c, e, and g) and fluorescent images (b, d, f, and h) of the transgenic lines at 36?48 hpf. Note the similar expression of GFP in the CNS in Wt, Del - 166/-146 and M4 constructs. Del - 1696/-166 exhibits no detectable GFP expression after 24 h post fertilization. In the fluorescent images there is variable auto-fluorescence in the yolk marked with an asterisk (*). (C) Transgenic fish harboring the Wt, Del-166/-146 or M4 mutant promoters driving GFP expression were harvested at 48 hpf. Retinal sections were stained with the nuclear dye Hoechst (a, e, and i), examined for GFP expression (b, f, and j) and stained with the Zn5 antibody that identifies differentiating RGCs (c, g, and k). Merged images showing GFP and Zn5 staining are also shown (d, h, and l). Note the Wt promoter-transgene is strongly expressed in the differentiating retinal ganglion cells, while Del - 166/-146 and M4 promoters exhibited a significant reduction in the number of GFP positive cells in the GCL. Three fish per line and two lines per transgene were assayed with similar results. (D) Transgenic fish harboring the Wt, Del-166/-146, M4 or Del-1696/-166 promoters driving GFP expression had their right optic nerve crushed on day 0. Six days later, left (control) and right (crush) retinas were isolated and assayed for endogenous tuba1a (α1Tubulin) mRNA by in situ hybridization (a, c, e, g, i, k, m, and o) or transgene promoter activity by GFP fluorescence (b, d, f, h, j, l, n, and p), respectively. In situ hybridization for tuba1a expression and GFP images are from adjacent retinal sections. At least three independent lines were assayed for each transgene, except M4 for which only two lines were identified. Three fish from each line were tested, all of which gave similar results. Note that although endogenous tuba1a (α1Tubulin) expression is highly induced following optic nerve crush in all fish tested only transgenic fish harboring the Wt tuba1a promoter show strong transgene GFP induction following optic nerve crush. White arrows indicate GFP positive RGCs, residing in the ganglion cell layer (GCL). |
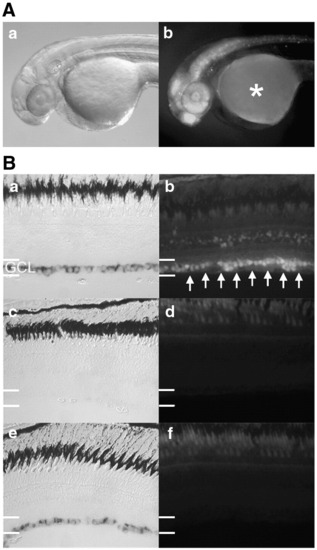
The Sp consensus binding site cannot functionally replace the G/C-rich enhancer in vivo. (A and B) Transgenic zebrafish were created which harbor the Sp consensus (SpCon) binding site replacing the G/C-rich enhancer in the tuba1a:GFP transgene. (A) SpCon transgenic fish display nervous system restricted GFP expression at 48 h post fertilization similar to Wt promoter (Fig. 8A). Auto-fluorescence in the yolk is noted with an asterisk (*). (B) SpCon transgenic fish fail to re-express GFP during optic nerve regeneration. Wt tuba1a:GFP transgenic fish exhibit strong tuba1a (a) and GFP (b) expression in the retinal ganglion cell layer (GCL) 3 days following optic nerve injury. In addition, we observed variable and weak GFP expression in inner nuclear layer cells in this particular line of fish which may reflect the transgene insertion site. Uninjured SpCon transgenic (c and d) and 3 day post-nerve injury (e and f) retinal sections were stained for tuba1a expression (c and e) or imaged for fluorescence (d and f). Note the absence of GFP fluorescence in the GCL of the injured retina (f) while endogenous tuba1a expression is highly induced (e). |
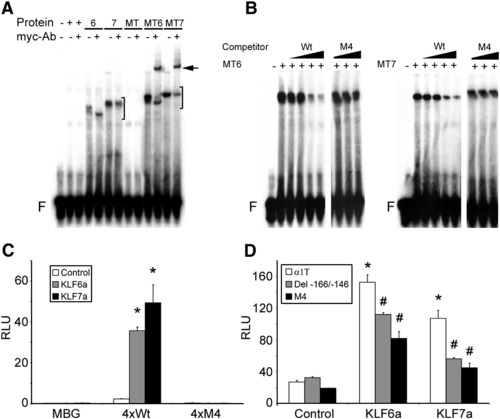
KLF6a and KLF7a bind the tuba1a G/C-rich element and can transactivate the tuba1a promoter through the G/C-rich element. (A) EMSAs were performed using radiolabeled G/C-rich element and in vitro synthesized KLF6a (6), KLF7a (7), myc-tag (MT), myc-tagged KLF6a (MT6), or myc-tagged KLF7a (MT7) protein. If no protein was added the lane is marked with a (-) and if the in vitro transcription translation reaction mix, lacking plasmid, was added the lane is marked with a (+). Both KLF6a and KLF7a bind to the probe (bracket) and in the cases where they harbor a myc-tag the anti-myc antibody (myc-Ab) results in a supershift of the complex (black arrow). (B) MT6 and MT7 binding is specific and can be completed by Wt oligonucleotide but not by M4 oligonucleotide (Wt and M4 EMSAs were performed on separate gels under identical conditions; M4 competitor concentrations are equal to the highest Wt competitor concentrations). Free probe (F). (C) KLF6a or KLF7a over expression can transactivate a heterologous promoter through the isolated G/C-rich element (*p < 0.01, t-test). (D) KLF6a or KLF7a overexpression can transactivate the tuba1a (α1T) promoter and this transactivation is attenuated when the G/C-rich element is deleted (Del - 166/-146) or mutated (M4) (*p < 0.01, ANOVA with Bonferonni post-hoc test comparing tuba1a (α1T) promoter driven expression with KLF6a or KLF7a overexpression, white bars) (#p < 0.05, ANOVA with Bonferroni post-hoc test comparing KLF6a or KLF7a overexpression effect on G/C-rich element deletion or mutation tuba1a (α1T):luciferase reporters, comparing grey or black bar to white bar). (C and D) PC12 cells were co-transfected with tuba1a (α1T):luciferase reporter vectors along with pCS2:β-gal and either empty pCS2 vector (Control) or pCS2-KLF6a (KLF6a) or pCS2-KLF7a (KLF7a). Reporter plasmids are: α1T, - 1696 tuba1a:luciferase; Del - 166/-146, internal deletion of the G/C-rich element in the - 1696 tuba1a promoter; M4, point mutant in G/C-rich element of - 1696 tuba1a promoter; MBG, minimal β-globin promoter; 4xWT, MBG with 4 copies of the G/C-rich enhancer cloned upstream; 4xM4, MBG with 4 copies of the M4 mutant G/C-rich enhancer cloned upstream. All transfections are normalized to pCS2:β-gal expression and luciferase values are reported as relative light units (RLU) in hundreds. Values are means +/- SEM. |
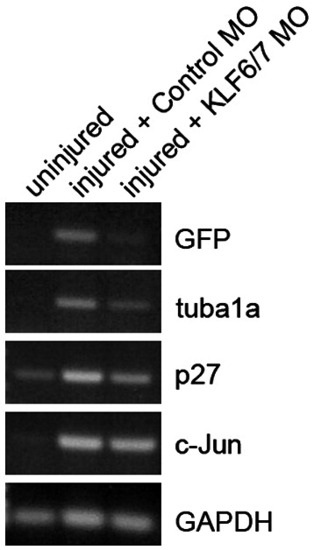
Regeneration-dependent induction of KLF6a and 7a is necessary for tuba1a and p27 gene induction following optic nerve injury. The optic nerve of adult tuba1a:GFP transgenic fish was lesioned and KLF6a/ KLF7a targeting MO or a control MO was applied to the optic nerve stump as described in Experimental methods. 3 days later retinas were harvested and RNA purified for use in RT-PCR reactions. Shown is an ethidium bromide stained gel showing induction of GFP, tuba1a, p27 and c-jun following optic nerve injury. GAPDH is used as a loading control. Note that KLF6a and 7a knockdown (KLF6/7 MO) caused a significant reduction in GFP, tuba1a and p27 gene induction following optic nerve injury. EXPRESSION / LABELING:
|

RGC axon regeneration is inhibited by tuba1a-targeting MOs in a concentration-dependent manner. Control or 2 different tuba1a-targeting MOs were delivered to RGCs, in vivo, by placing MO-soaked Gelfoam on the sectioned optic nerve stump. The next day the Gelfoam was removed and 3 days later retinas were isolated, diced and placed in explant culture for 4 days prior to analysis of neurite outgrowth. The graph on the right shows the Nerve Growth Index (see Experimental methods) for each MO at the concentration indicated. Note the concentration-dependent reduction in neurite outgrowth (Nerve Growth Index) as increasing concentrations of the tuba1a-targeting MO are applied to the optic nerve stump. Also note that tuba1a-targeting MOs #1 and #2 at 0.065 mM concentration have little effect on neurite outgrowth when applied individually, but synergize to significantly reduce neurite outgrowth when added together. The image on the left shows representative explants treated with tuba1a-targeting MOs #1 and/or #2 at 0.065 mM concentration. Note the reduced axon outgrowth in explants from retinas that received a combination of MOs #1 and #2. |
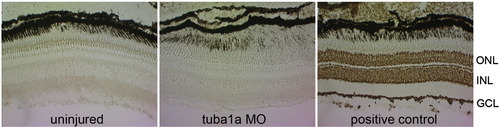
Tuba1a-targeting MOs do not stimulate RGC apoptosis. tuba1a-targeting MO was delivered to RGCs, in vivo, by placing MO-soaked Gelfoam on the sectioned optic nerve stump. The next day the Gelfoam was removed and 3 days later retinas were isolated, sectioned and assayed for cell death using TUNEL stain. Note that like the uninjured retina, MO-treated retinas showed little cell death. Positive control is a nuclease-treated retina. Brown signal indicates cells undergoing apoptosis. |
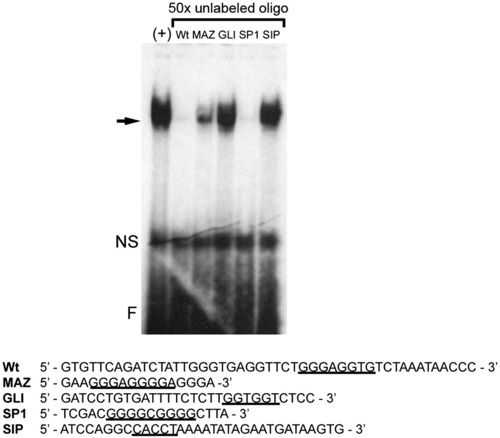
The Sp1 consensus sequence can compete for nuclear protein binding to the G/C-rich enhancer. Electrophoretic mobility shift assay showing the Sp1 consensus binding site probe can efficiently compete nuclear protein binding to the G/C-rich enhancer. Below the gel are shown the oligonucleotide sequences of consensus protein binding sites for myc-associated zinc finger protein (MAZ), GLI, specificity protein 1 (SP1), and smad interacting protein (SIP) used as unlabeled competitors. Consensus binding sites are underlined. The arrow marks specific protein binding. NS, Nonspecific binding; F, free probe. |
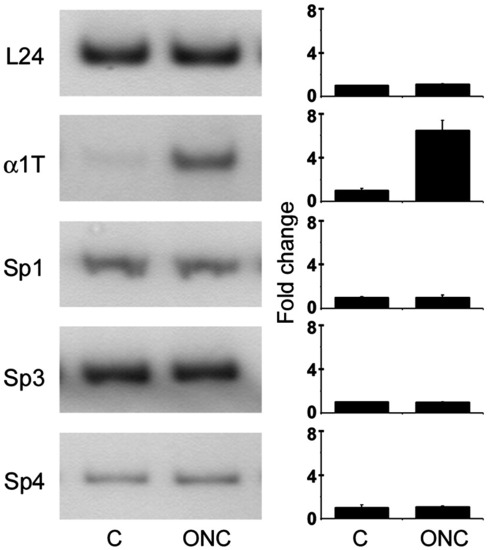
Sp proteins are not induced during optic nerve regeneration. Sp1, Sp3, and Sp4 expressions are not changed during optic nerve regeneration in zebrafish. Quantitative RT-PCR for negative control ribosomal protein encoding transcript L24, positive control tuba1a (α1T), and Sp1, Sp3, and Sp4 using uninjured retinal RNA and 3 day post-optic nerve crush retinal RNA (ONC) as template. Bar graphs represent densitometric quantification of autoradiographic analysis of RT-PCR products from 3 separate fish. |
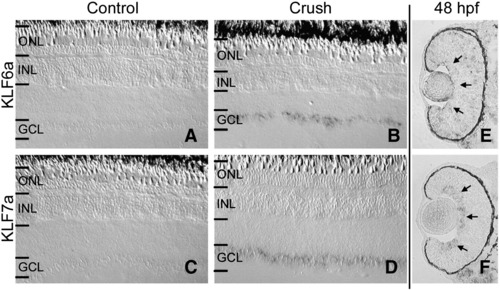
KLF6a and KLF7a are induced in regenerating RGCs following optic nerve injury, but only KLF7a is expressed in differentiating RGCs. (A and B) In situ hybridization showing KLF6a induction in retinal ganglion cells 3 days post-optic nerve crush (B) as compared to uninjured retina (A). (C and D) In situ hybridization showing KLF7a induction in retinal ganglion cells 3 days post-optic nerve crush (D) as compared to uninjured retina (C). (E) KLF6a is not expressed in the retina at 48 h post fertilization (hpf). (F) KLF7a is expressed in the differentiating RGCs at 48 hpf. Arrows denote developing GCL. Outer nuclear layer (OLN), inner nuclear layer (INL), and ganglion cell layer (GCL). |
Reprinted from Molecular and cellular neurosciences, 43(4), Veldman, M.B., Bemben, M.A., and Goldman, D., Tuba1a gene expression is regulated by KLF6/7 and is necessary for CNS development and regeneration in zebrafish, 370-383, Copyright (2010) with permission from Elsevier. Full text @ Mol. Cell Neurosci.