|
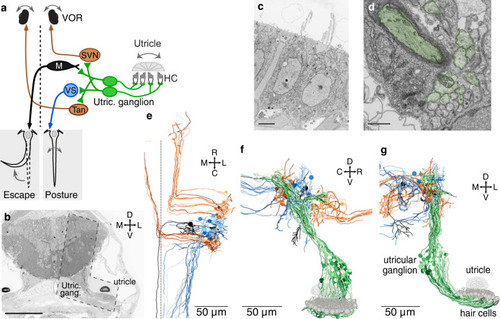
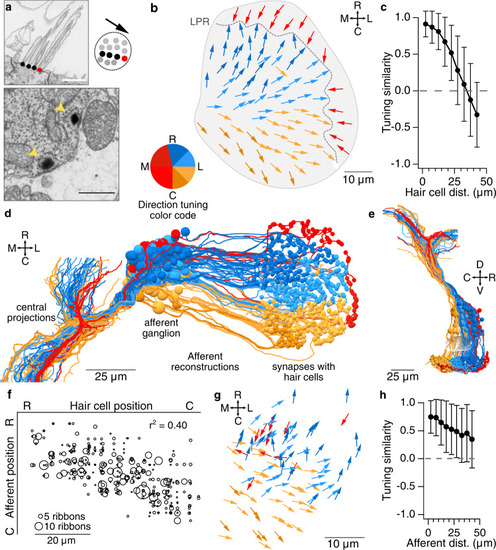
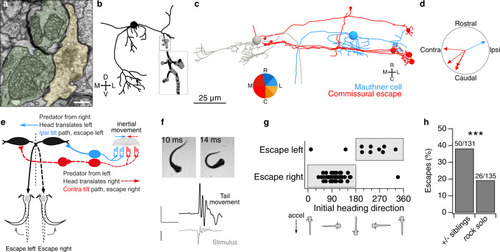
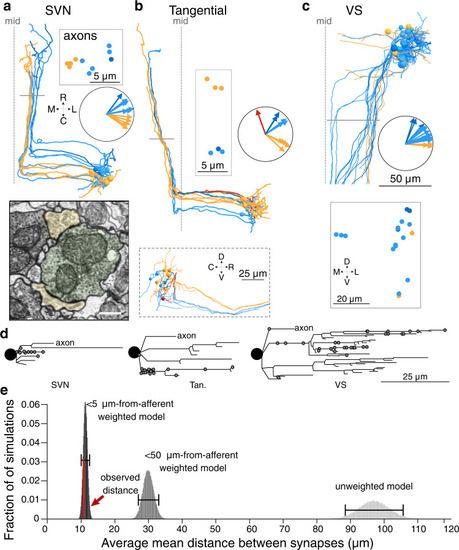
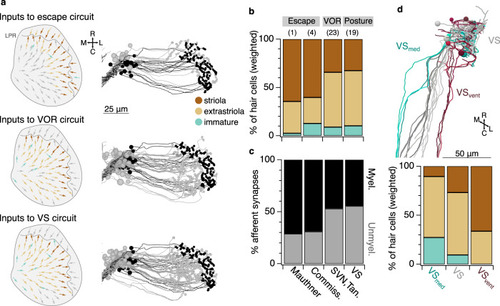
Early developing afferent pathways with fast kinetics preferentially drive early developing central neurons for fast escapes.a Hair cells in the utricular macula that excite afferents connected to Mauthner escape, VOR, or VS circuits (top, middle, and bottom). Hair cells are colorized if they contribute input to a pathway or gray if they do not. Right, afferents connected to escape, VOR, or VS circuits, colorized by their myelination status (black: myelinated; gray, unmyelinated). b Quantification of the contribution of striolar, extrastriolar, and immature hair cells to these central pathways. Striolar hair cells preferentially drive rapid escape circuits, both via the direct afferent input to the ipsilateral Mauthner and the afferent input to the commissural escape pathway, whereas VOR and VS circuits receive input from a mixture of pathways, with extrastriolar inputs dominating. Numbers in parentheses indicate the number of central neurons in each category. Wilcoxon test, striolar contribution to escape vs non-escape neurons, p = 0.041. c Quantification of the contribution of synapses from myelinated and unmyelinated afferents to each central pathway. Afferents driving escape pathways are largely myelinated at this age, whereas afferents driving VOR and postural pathways are more mixed. Chi-square test for all groups, p = 6.0 × 10−8. Follow-up chi-square: VOR vs escape, 1.4 × 10−4; VS vs escape, 1.7 × 10−8. d Subsets of VS neurons, identified by axon trajectories, are predicted to exhibit different temporal kinetics. VS neurons with axons that approach the midline before descending (VSmed, greens) receive mostly extrastriolar (tonic) input with a large contribution from immature hair cells, whereas VS neurons with ventral axon trajectories (VSvent, dark reds) receive mostly striolar (phasic) input and none from immature pathways. The skeleton reconstruction is projected at a mixed horizontal/sagittal angle to facilitate visualization of these groups. See also representation of axon trajectories in Fig. 4c and Supplementary Fig. 4.
|