
- Title
-
Wnt9 directs zebrafish heart tube assembly via a combination of canonical and non-canonical pathway signaling
- Authors
- Paolini, A., Sharipova, D., Lange, T., Abdelilah-Seyfried, S.
- Source
- Full text @ Development
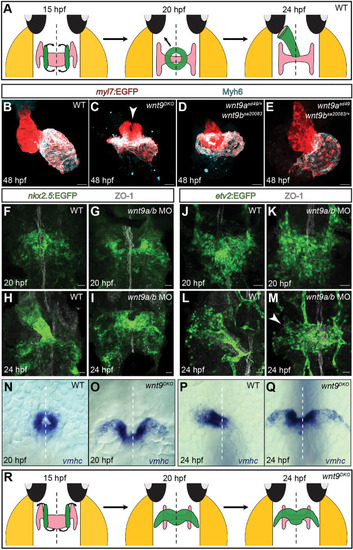
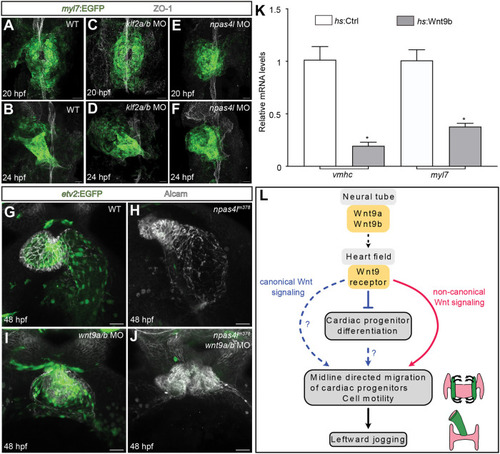
EXPRESSION / LABELING:
PHENOTYPE:
|
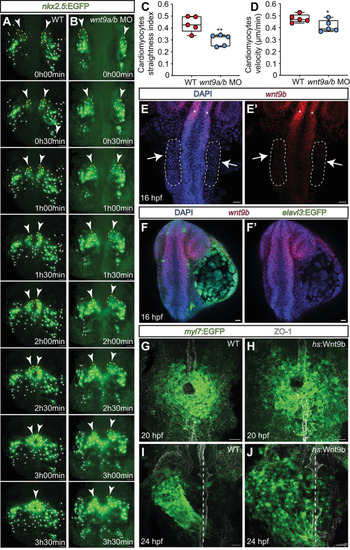
EXPRESSION / LABELING:
PHENOTYPE:
|
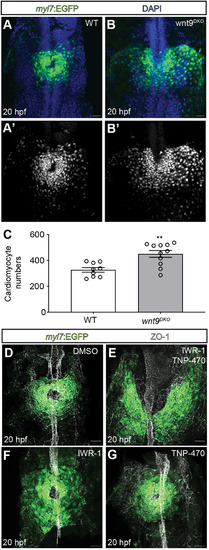
EXPRESSION / LABELING:
PHENOTYPE:
|
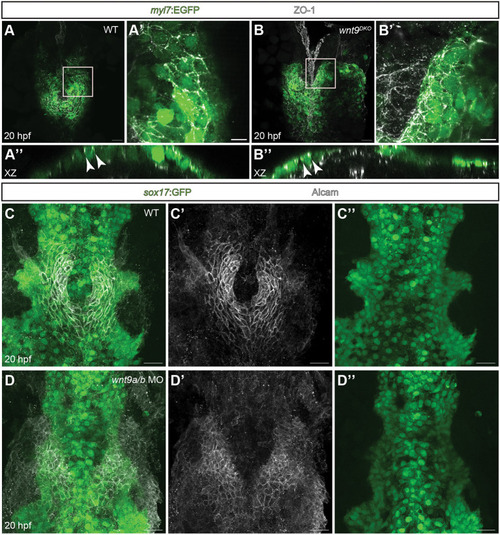
EXPRESSION / LABELING:
PHENOTYPE:
|
EXPRESSION / LABELING:
PHENOTYPE:
|