
- Title
-
Re-epithelialization of cutaneous wounds in adult zebrafish uses a combination of mechanisms at play during wound closure in embryonic and adult mammals
- Authors
- Richardson, R., Metzger, M., Knyphausen, P., Ramezani, T., Slanchev, K., Kraus, C., Schmelzer, E., Hammerschmidt, M.
- Source
- Full text @ Development
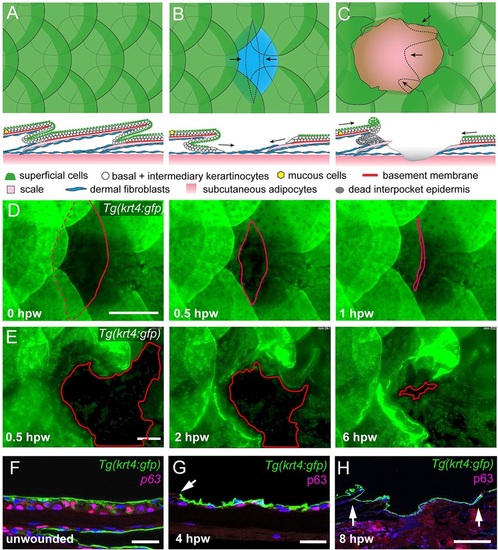
Re-epithelialization of full- and partial-thickness wounds in adult zebrafish. (A-C) Schematic representations of normal adult zebrafish skin architecture (A), or following partial-thickness (B) or full-thickness (C) wound introduction. Arrows in B and C indicate the typical directions of epidermal re-epithelialization. (D,E) Single images from time-lapse recordings of Tg(krt4:GFP) fish following partial-thickness (D) or full-thickness (E) wounding at indicated time points post-wounding; red line marks LE. (F-H) Unwounded epidermis is 3-4 cell layers thick (F), the re-epithelializing neo-epidermis is bilayered, consisting of superficial GFP+ and inner p63+ keratinocytes (G,H). Arrow in G indicates LE, arrows in H, the wound margins, revealing full re-epithelialization at 8hpw. Scale bars: 500�m in D,E,H; 20�m in F,G. EXPRESSION / LABELING:
|

LE cells of partial-thickness wounds form lamellipodia but lack signs of purse-string mechanisms. (A) Puncture wound in fin of 3-day-old embryo shows circular, colocalized actin (left panel) and phosphorylated non-muscle myosin (middle panel) at the wound margin. Images are maximum projections, insets single z-planes in the centre of the wound. (B) Puncture wound in trunk of adult, with cortical p-myosin in adjacent keratinocytes, including the sides facing the wound (arrow). (C,D) Adult partial-thickness wound; actin fibres are present within the protruding lamellipodia (red), p-myosin is accumulated at the posterior of LE cells (magenta, arrow). (E-G) Live confocal images of partial-thickness wound of Tg(actb2:hras-egfp), Tg(krt4:mCherry) double transgenic. Lamellipodia are formed at the LE by inner keratinocytes (green), whereas superficial cells (red) remain further back (E,F). (G) Single plane confocal images, revealing the enormous lamellipodial size relative to the cell body of an LE keratinocyte (upper panel). Inner keratinocytes behind the LE also show restricted protrusive activity (G, bottom panel; lamellipodium, arrow). (H) Stills from a time-lapse movie at indicated relative time points. Rarely, usually when encountering an obstruction, LE cells undergo more pronounced EMT-like changes and retain only limited contact with following cells. Asterisks indicate the same blood vessel. Scale bars: 20�m in A-H; 10�m in insets in A. EXPRESSION / LABELING:
|
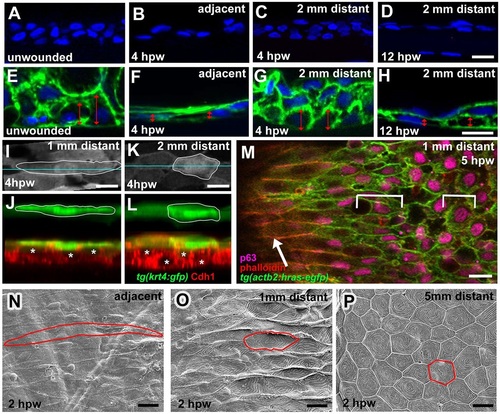
Epidermal cells outside full-thickness wounds undergo progressive radial intercalation, flattening and elongation. (A-D) DAPI labelling of sections through unwounded epidermis (A) and epidermis at indicated stages and distances from wound (B-D). (E-H) DAPI labelling of sections through Tg(actb2:hras-egfp) fish as in A-D. Cell membranes are in green. Doubled-headed red arrows indicate the heights of individual basal keratinocytes. (I-L) Surface views (I,K) and z-projections (J,L) of Tg(krt4:GFP) fish; at 1mm distance, superficial cells (outlined; green) display more pronounced flattening, and inner keratinocytes (E-cadherin; red; centres indicated by asterisks) more pronounced radial intercalations than at 2mm distance from wound. (M) Single-plane confocal micrograph of Tg(actb2:hras-egfp) fish; superficial cells are labelled with Phalloidin (red), inner epidermal cells with p63 (pink) and cell membranes with GFP (green); wound is to the left. Arrow indicates superficial cells, brackets indicate lengths of inner keratinocytes, which are less elongated than superficial cells. (N-P) SEM images of superficial skin layer (single cells outlined) at indicated distances from wound, revealing spatially graded cell elongation and loss of surface microridges. Scale bars: 10�m. EXPRESSION / LABELING:
|

Re-epithelialization requires TGF-β/GRGDS-dependent keratinocyte crawling at the LE and Rock/JNK-dependent epithelial morphogenesis in the following epidermis. (A-D) Quantification of effects caused by the indicated inhibitors on LE migration speeds in partial-thickness wounds (A), and on the extent of wound closure (B), the degree of circularity (C) and the orientation of the long axis (D) of adjacent superficial cells of full-thickness wounds at 10hpw. Tg(DEP), transgenic inhibition of PCP. In A-C, mean values and s.d. are indicated. Values in A were determined from time-lapse movies (e.g. Movies 2-5; n≥5 per condition), values in B from Methylene Blue penetration assays as shown in Fig. S4F,G, and values in C,D from images as shown in O-S. *P<0.05; **P<0.01; ***P<0.005; ****P<0.001; n.s., not significant; one-way ANOVA with a Dunnett′s post-hoc test. (E-N) Live confocal images of the LE of Tg(actb2:hras-egfp), Tg(krt4:mCherry) double transgenic fish at 5min (E-I) and 30min (J-N) after partial-thickness wounding (mpw), treated with indicated inhibitors. Lower panels of J-N show z-projections of the LE cell marked by the asterisk in the panels above. At 5mpw, all cases except the CYT-D treatment display normal protrusive activity at the LE, whereas at 30mpw, SB431542- and GRGDS-treated LE keratinocytes display roundish, shorter and thicker lamellipodia, indicative of lamellipodial retraction. (O-S) Phalloidin staining of the adjacent epidermis of full-thickness wounds at 10hpw demonstrating compromised and uncoordinated cell elongation upon treatment with CYT-D (P) and Y27632 (S), but not SB431542 (Q) or GRGDS (S). (T-V) p63, GFP double immunofluorescence and DAPI labelling of sections through Tg(actb2:hras-egfp) fish at indicated conditions and distances from full-thickness wounds. Left panels, merge; right panels, p63 channels. Y27632-treated wound (V) displays reduced flattening of superficial cells (double-headed red arrows) and reduced frequency of radial intercalations, characterized by a partial overlap of the positions along the epidermal apical-basal axis occupied by adjacent basal and suprabasal p63+ nuclei (marked in U by red lines). Quantifications from three individual fish per condition (1-1.5mm wound distance for U,V) yielded the following frequencies (no. of partially overlapping p63+ nuclei/total no. of p63+ nuclei). Unwounded in T, 6.1�5.3%; DMSO-treated wound in U, 51.3�3.7%; Y27632-treated wound in V, 17.2�4.9%. P=0.00027557 (T versus U); P=0.056567873 (T versus V); P=0.000651499 (U versus V); Student′s t-test. Scale bars: 20�m. EXPRESSION / LABELING:
|

Epidermal cell proliferation regenerates the surrounding epidermis but does not influence re-epithelialization. (A-C) Histological analysis reveals a highly thickened neo-epidermis at both 24 and 72hpw (A). The epidermis 1mm (B) and 2mm (C) distant to the wound is much thinner at 24hpw, but has recovered to normal thickness by 72hpw. (D-I) BrdU-incorporation studies reveal few labelled cells in the wound epidermis from 24hpw to 72hpw (D-F), but a strong increase of BrdU+ cells in the epidermis 1mm distant from the wound (G-I). (J-O) Fish treated with hydroxyurea (HU) exhibit the same rate of re-epithelialization as assessed by Methylene Blue assay at 8hpw (J,K). Histological analysis between control and HU-treated fish reveals similar epidermal thicknesses in the wound at 24hpw and 72hpw (L,M), and 1mm distant from the wound at 24hpw (N,O). However, at 72hpw, the latter has recovered to its normal thickness in the control, but remains thin in the HU-treated fish (N,O). Scale bars: 20�m in A-C, L-O; 50�m in D-J. |
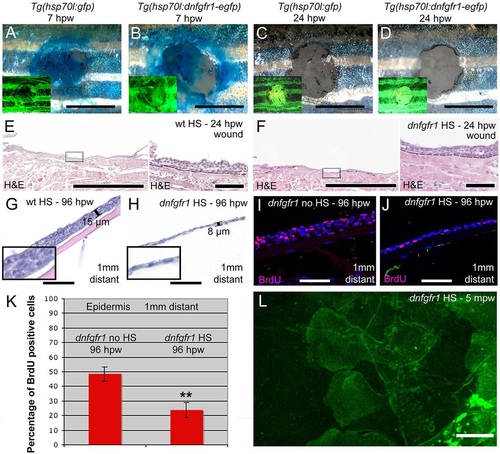
FGF signalling is dispensable for re-epithelialization, but is required for keratinocyte proliferation and epidermal regrowth. (A-D) Methylene Blue penetration assay at 7hpw (A,B) and 24hpw (C,D) revealing unaltered re-epithelialization rates of full-thickness wounds between heat-shocked control Tg(hsp70l:gfp) (A,C) and Tg(hsp70l:dnfgfr1-egfp) (B,D). Insets show fluorescence of transgene-encoded GFP or dnFGFR-GFP fusion proteins, indicating strong transgene expression. (E,F) Histological analysis of wound epidermis at 24hpw revealing unaltered thicknesses of neo-epidermis between heat-shocked wild-type control (E) and Tg(hsp70l:dnfgfr1-egfp) fish (F). Right panels show magnified views of regions boxed in left panels. (G,H) At 96hpw, the epidermis 1mm distant from the wound of the heat-shocked Tg(hsp70l:dnfgfr1-egfp) fish (H) is much thinner than in the heat-shocked non-transgenic control (G). Insets show magnified views. (I,J) BrdU incorporation revealing significantly reduced epidermal proliferation in heat-shocked (J) compared with non-heat shocked Tg(hsp70l:dnfgfr1-egfp) fish (I) 1mm distant from the wound. (K) Quantification of BrdU incorporation rates (percentage of cells) from images as in I,J. (L) Live image of LE of heat-shocked Tg(hsp70l:dnfgfr1-egfp) fish after partial-thickness injury, revealing normal protrusive activity (see Fig. 4E as control). Scale bars: 2mm in A-D; 500�m in left panels of E,F; 50�m in G-J and right panels of E,F; 10�m in L. EXPRESSION / LABELING:
|
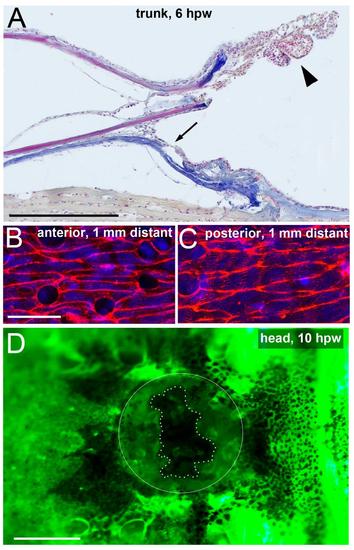
Keratinocytes from both anterior and posterior regions move towards full-thickness wounds. Longitudinal section through a full-thickness wound at 6 hpw, stained with AFOG; only the anterior part of the wound is shown; anterior to the right. Arrow points to anterior border of neo-epidermis formed via re-epithelialization from the posterior side. Arrowhead points to pile of keratinocytes at tip of anterior scale remnant, which have largely lost direct contact to the scale surface (compare with Supplementary Movie 1 and Fig. 1C). (B,C) Phalloidin staining of superficial keratinocytes in regions 1 mm anterior (B) and 1 mm posterior (C) from a full-thickness wound, 4 hpw. Note that on both sides, keratinocytes have elongated in a directed manner (compare with Fig. 4P as negative control). The elongation in anterior regions indicates that these cellular arrangements are largely independent of ?pulling? from the LE, as cells pile up at the tip of the scale remnant (see A), rather than re-populating the wound bed. (D) Live fluorescent image of a Tg(krt4:GFP) fish after introduction of a full-thickness wound on the forehead; 10 hpw; anterior to the left; wound encircled; edges of neoepidermis marked by dotted line. In contrast to trunk wounds (compare with Movie 1), the head wound is re-epithelialized from all sides, including the anterior side. Closure of shown wound: 70%; average (n=6): 73�7.7%. Scale bars: A,D: 1 mm; B,C: 20 �m. |
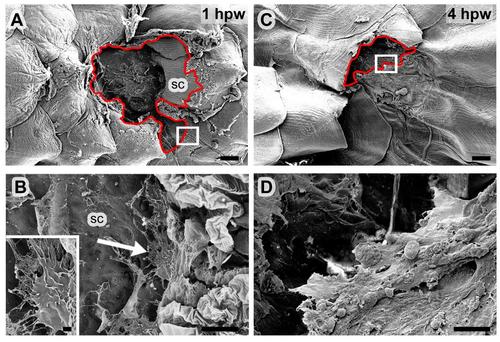
LE keratinocytes of full-thickness wounds only display protrusive activity when positioned on naked scale remnants. All panels show SEM micrographs of 1 mm full-thickness wounds; (B,D) show magnified views of regions boxed in (A,C), inset in (B) shows magnified view of cellular protrusion indicated by arrow in (B). In (A,C), wound edges / leading edges of epidermal tongues are marked by red lines. Leading edge cells only extend protrusions in regions where the substrate allows, for example, on remnants of a damaged scale (A,B). In contrast, re-epithelializing epidermis covering the denuded muscle layer at the centre of the wound does not seem to be directly attached to a migration substrate, and the LE cells do not show distinct protrusions (C,D). Abbreviation: sc, scale. Scale bars: A,C = 200 �m; B,D = 10 �m; inset in B = 1 �m. |

The epidermis around closing full-thickness wounds lacks apoptotic keratinocytes. (A-C) TUNEL staining on sections through unwounded (A.B) and wounded (C) trunk region, 4 hpw, with (B) and without (A,C) DNase treatment to induce DNA fragmentation, as formerly described (Fischer et al., 2014). No TUNEL-positive cells are detectable in the epidermis 1 mm from the wound, indicating that the thinning of the epidermis in this region is not due to keratinocyte apoptosis. Similar results were obtained via immunofluorescent labelling of activated caspase 3 (data not shown). Abbreviation: mc, melanocyte. |
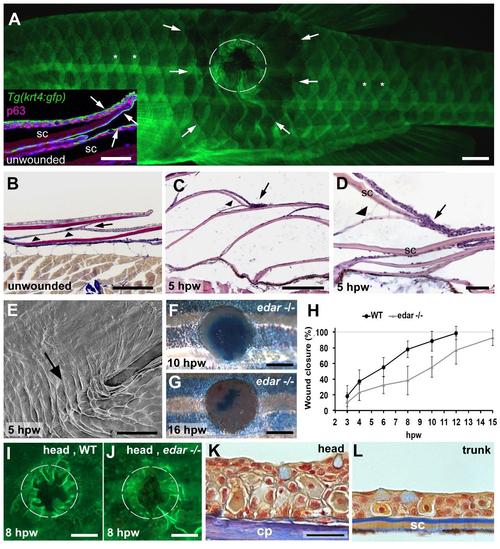
Re-epithelialization uses epidermal inter-scale reservoirs. (A) Overview of a Tg(krt4:gfp) wounded fish at 5 hpw. In unwounded regions, the posterior tip of the scales exhibit stronger GFP signal (marked by asterisks) due to overlapping GFP-positive cells wrapping the scale tip (arrowed in immunofluorescence of sections in inset). In a large area around the wound, the epidermis appears less GFP-positive (arrows depict the outer margin). The circle marks the approximate extent of the wound. (B-D) Histological analysis of the unwounded skin (B) and region adjacent to the wound (C,D), demonstrating the bridging of the epidermis from one scale to the next (indicated by arrows). Panel (D) shows higher magnification of such a ?bridge?, with epidermal cells on the apical surface of one scale crossing onto the apical surface of the adjacent scale with no wrapping of the posterior tip of the scale. The only cellular structure seen beneath the scale is the dermal tether (indicated by arrowheads). (E) SEM micrograph of epidermal surface adjacent to the wound. The bridging epidermis between two adjacent scales is indicated by the arrow. (F,G) Methylene blue analysis of full thickness wounds to the flank of edar-/- fish indicates that these wounds re-epithelialize slower than in wild type and are not fully closed at 16 hpw. (H) Graphical illustration of the kinetics of wound closure of edar-/- and wild-type control fish, determined via the methylene blue penetration assay at indicated time points after wounding. Mean values and standard deviations are indicated. (I,J) Live fluorescent images of Tg(krt4:GFP) wild-type (I) and edar-/- (J) fish after introduction of full-thickness wounds on the forehead; 8 hpw; anterior to the left; wound encircled. Head wounds of wild-type fish (I: 63%; average: 65�14.1%; n=11) and edar mutants (J: 63%; average: 65�14.1%; n=9) display similar degrees of closure. However, both are less closed than equally-sized 8 hpw wounds in the flank of wild-type fish (78�10.6%; 6 fish; panel H), and more closed than equally-sized 8 hpw in the flank of edar mutants (38�18.0%; 6 fish; panel H). (K,L) AFOG staining of longitudinal section through head (K) and trunk (L) of 1-year old unwounded wild-type fish; anterior to the left. The epidermis on the forehead consists of 6-8 keratinocyte layers and is approximately 300 �m thick, the epidermis on the scales of the trunk consists of 3-4 keratinocyte layers and is approximately 150 �m thick. Since the head epidermis sits almost directly on the bony calvarial plates of the skull (K), reduced laser settings had to be used for wounding (specified in Materials & Methods) to avoid damage to the skull. Abbreviations: cp, calvarial plate of skull; sc, scale. Scale bars: A,F,G,I,J = 1mm; D,E, inset in A = 50 �m; B,C,K,L = 200 �m. Due to the different laser settings during wound introduction, direct comparisons of wild-type wounds in the (scale-bearing) trunk and the (scale-free) head (H,I) with respect to the impact of inter-scale pockets have to be taken with caution. The same applies to direct comparisons of trunk wounds in (scale-bearing) wild-type fish and (scale-free) edar mutants (H), as during scale placode formation, edar mutants also present defects in epidermal cells themselves (Harris et al., 2008). Thus, the lack of inter-scale reservoirs may not be the only cause of the wound closure defect. However, it is noteworthy that wounds in scale-free head regions lacking the inter-scale pockets close more slowly than wild-type trunk wounds (H,I), although the higher thickness of the head epidermis (K,L) should have a positive effect on re-epithelialization, as it contains more keratinocytes that could be recruited during wound re-epithelialization. Consistent with the latter notion, edar mutants, which lack scales and inter-scale pockets both in head and trunk, head wounds close faster than trunk wounds (F,J). Together, this points to a more efficient recruitment of keratinocytes from inter-scale pockets than from an accordingly thicker epidermis. Furthermore, the unaltered closure rates of head wounds of edar mutants compared to wild-type siblings (I,J) strongly argues against a general negative effect of the edar mutation on the re-epithelializing capabilities of keratinocytes, suggesting that the reduced closure rates of trunk wounds in edar mutants compared to wild-type fish (H) are indeed primarily caused by the absence of the inter-scale pockets. |

Efficacy of SB-431542 (TGFβ) and SP600125 (JNK) inhibitors. (A,B) Live images of embryos treated with 0.1% DMSO (A, control) or with 10 �M SB431542 (B), starting at 4 hours post fertilization (hpf); 24 hpf, lateral view. The SB431542-treated embryo displays fused eyes (arrowhead) and lack of trunk somites (arrow), resembling the phenotype of mutants lacking the TGFβ family members Ndr1 and Ndr2 (Feldman et al., 1999), indicating that 10 �M SB431542 efficiently blocks TGFβ signalling (n=45/48; 2 independent experiments). (C) Duplex anti-pJNK (1:1000, Cell Signaling Technology, 4668s) and anti-&beta-actin (loading control; 1:1000, Sigma-Aldrich, A2228) Western blot of protein extracts from fins of adult zebrafish, after treatment with 0.1% DMSO (control) or 10 �M SP600125 for 5 hours, and separation via 9% SDS-PAGE. Levels of the phosphorylated version of the p54 splice isoform of JNK are several-fold reduced in the SP600125-treated sample. Similar results were obtained in three independent experiments. |
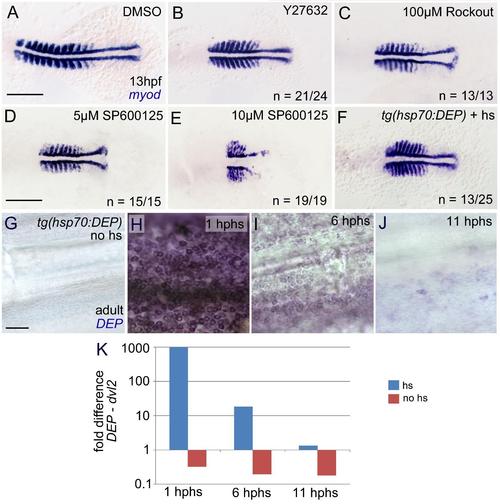
The Rock inhibitors Y27632 and Rockout and the JNK inhibitor SP600125 cause compromised convergent extension similar to the effects of transgenic inhibition of the PCP pathway.(A-F) Whole mount myoD in situ hybridization of presumptive paraxial muscle cells at 13 hpf (10-somite stage); dorsal views on flat-mounts; anterior to the left. Embryos shown in (A,C-E) had been incubated in medium containing the indicated drugs at the indicated concentration, starting at 4 hpf; embryo shown in (B) had been microinjected with Y-27632 at the 1-cell stage (2 nl of 400 �M solution), as described (Lai et al., 2005; Lou et al., 2012). Therefore, no final Y-27632 concentration is supplied in the panel. However, assuming that the cytoplasm of freshly laid eggs has a volume of 0.6 mm3 (1/3 of the total volume of the egg), it should be approximately 1 �M). Embryo shown in (F) derives from a cross of Tg(hsp70l:DEP) transgenic and a wild-type parent, and was subjected to a 40�C heat shock from 4 to 4.5 hpf. Only approximately 50% of the offspring were affected, corresponding to the expected ratio of transgene segregation. Compared to the control (A), embryos shown in (B-F) display a shorter and broader body axis, similar to mutants in PCP pathway components (Marlow et al., 2002; and references therein). (G-J) Xenopus Dishevelled DEP in situ hybridization of amputated tail fins from Tg(hsp70l:DEP) adults without heat shock (G) and at 1 hour, 6 hours and 11 hours after heat shock application (hphs). Transcripts encoding the DEP domain of Xenopus Dishevelled are readily detectable at 1 (H) and, at significantly lower levels, at 6 (I) and 11 hphs (J). This points to efficient transgene expression, but limited stability of the transgene-encoded transcripts. (K) Xenopus Dishevelled DEP qRT-PCR analysis of RNA extracted from the skin of the wounded flank of heat-shocked (blue) or, as control, non-heat-shocked (red) Tg(hsp70l:DEP) adults with a 7500 Fast Real Time PCR System (Applied Biosystems, Waltham, MA) and SYBR green for DNA quantification. Obtained values were normalized against gapdh and compared to endogenous dvl2 at three different time points after heatshock application and full-thickness wounding: 1 hour after heat shock (hphs), 0 hpw: 987-fold higher; 6 hphs, 5 hpw: 18-fold higher; 11 hphs, 10 hpw: 1.3-fold higher expression levels of transgenic dsh-DEP compared to endogenous dvl2. Sequences of primers used were: Xenopus dsh-DEP forward: TTGTGAAGGTCATGGCTTCC, reverse: ATAGAGCCAGTCCACCACAT; zebrafish dvl2 forward: GCCTCCTCTTTCAGCAGTGT, reverse: CCCACGATACTGATGCCCAA; gapdh forward: CGCTGGCATCTCCCTCAA, reverse: TCAGCAACACGATGGCTGTAG. Similar results were obtained in three independent experiments. For wound closure analyses, transgenic fish were either heat-shocked once (1 hour before wounding; see Materials & Methods) or twice (1 hour before and 5 hours after wounding), yielding indistinguishable results (insignificantly reduced closure rates; Fig. 4B and data not shown). This suggests that a single heatshock application is sufficient to block PCP signaling, and provides further evidence that PCP signaling is largely dispensable for wound closure. Scale bars: A-F: 200 �m; G,H: 50 �m. |
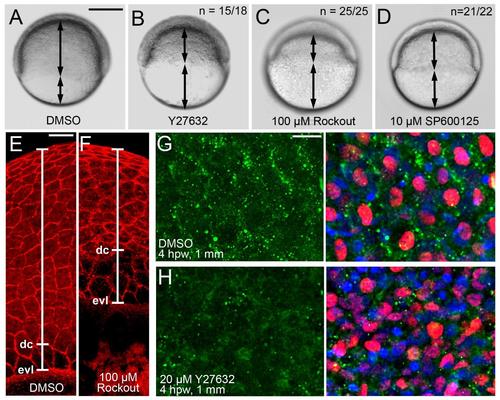
Chemical inhibition of Rock or JNK leads to compromised epiboly movements in gastrulating embryos and to reduced p-myosin II levels in the epidermis around full-thickness wounds of adult zebrafish. (A-D) Live images of embryos at 7 hpw (70% epiboly stage for control; A). Embryos shown in (A,C,D) had been incubated in medium containing the indicated drugs at the indicated concentration, starting at 4 hpf; embryo shown in (B) had been microinjected with Y-27632 at the 1-cell stage (2 nl of 400 �M solution). Regions covered by deep cells and uncovered yolk are indicated by arrows. The outer enveloping layer (EVL) is not visible in these images. Rock and JNK inhibition lead to compromised deep cell epiboly. (E,F) Embryos as shown in (A,C), after phalloidin staining of actin cytoskeleton. In addition to deep cells, Rock inhibition also leads to delayed epiboly of the EVL, possibly pointing to an essential role of Rock during EVL cell flattening, which is a driver a EVL epiboly (Slanchev et al., 2009). However, deep cell (dc) epiboly is more severely compromised than EVL (evl) epiboly, indicated by the increased distance between the margins of the two tissues on the yolk (marked by white line). These differential and dissectable effects point to an additional role of Rock during radial intercalation, which is a major driving force of deep cell, but not EVL epiboly, consistent with results obtained in former mutant analyses (Slanchev et al., 2009). (G,H) Confocal images of epidermis at 1 mm distance from full-thickness wound of adults, 4 hpw, immunolabeled for phosphorylated non-muscle myosin (green) and p63 (red), and counterstained with DAPI (blue). Left panels show pMyosin channel, right panels merged images. p63+ keratinocytes of the DMSO-treated control fish display stronger pMyosin labelling particularly in cortical regions (G), while labelling in the Y27632-treated fish is much reduced and less localized (H). The punctate localization in (G) is in line with the reported punctate actin contractions taking place during morphogenetic movements in Xenopus embryos (Kim and Davidson, 2010). Higher nuclei density in (H) is a reflection of the compromised thinning of the epidermis. Scale bars: A-D: 200 �m; E,F: 50 �m; G,H: 10 �m. |