
- Title
-
Ploidy manipulation and induction of alternate cleavage patterns through inhibition of centrosome duplication in the early zebrafish embryo
- Authors
- Heier, J., Takle, K.A., Hasley, A.O., Pelegri, F.
- Source
- Full text @ Dev. Dyn.
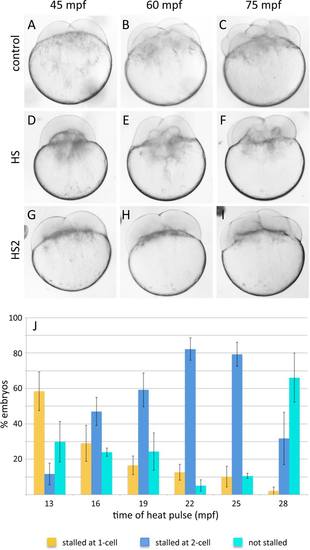
Standard Heat Shock causes a cell cleavage stall at the first cell cycle, and HS2 a stall at the second cell cycle. A?C: Control untreated embryos at the indicated time points, corresponding to the two-, four-, and eight-cell stage. D?F: An embryo with a one cell cycle stall during the first cell division, caused by a heat pulse initiated at 13 mpf (standard Heat Shock (HS)). G?I: An embryo with a one cell cycle stall during the second cell division, caused by a heat pulse initiated at 22 mpf (HS2). Embryos exhibiting a cell cycle stall resume cleavage after the stall and contain half the number of blastomeres as control stage-matched embryos. J: Comparison of the degree and type of cell cycle stall caused by standard Heat Shock and HS2. Standard Heat Shock causes primarily a stall of the first cell division, whereas HS2 primarily causes a stall of the second cell division. Values are mean percentages across four different trials. |
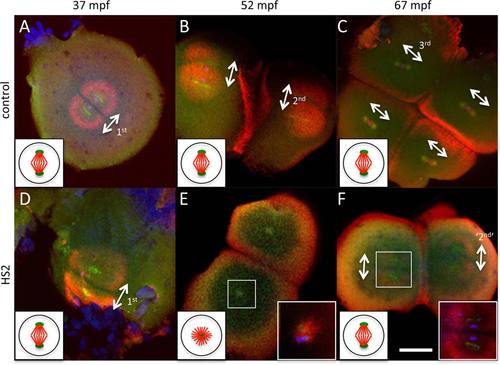
HS2 causes defects in bipolar spindle formation during the second cell cycle, likely caused by centrosomal duplication defects during the first cycle. Immunofluorescence microscopy of whole mount blastodiscs, labeled to detect α-tubulin (red), γ-tubulin (green), and DNA (blue, only readily apparent in high magnification inserts in E,F) in control (A?C) and HS2-treated (D?F) embryos. Bipolar spindle is normal during the first cell division cycle (A,D). During the second cell division cycle, each of the two previously generated blastomeres show monoasters in HS2-treated embryos (E, insert) instead of bipolar spindles in control embryos (B). One cycle later, HS2-treated embryos resume the formation of bipolar spindles and cell cleavage (F, insert), although the number of blastomeres is halved compared with control embryos (forming four blastomeres in HS2-treated embryos [F] and eight in control embryos [C]). Centrosomal material can be observed near the center of astral structures in both bipolar spindles and monoasters. In all cells, this centrosomal material appears duplicated with centers at each of the spindle poles, except in HS2-treated embryos during the second cell cycle, where centrosomal material appears as a single mass at the center of a monoaster (E, insert). The absence of a pair of centrosomes during the second cell cycle in HS2-treated embryos is presumably caused by defects in centriolar duplication during the first cell cycle, the time of heat treatment. The bipolar spindle is a dynamic structure that appears first, during metaphase, as a tight array of midzone microtubules (as in C) and later, during anaphase, as a diastral structure (A,B,D,F). Where present, white double-arrows indicate axes of bipolar spindles for the corresponding cell cycle (1st, 2nd, or 3rd, with ?2nd? indicating the bipolar spindle that forms after the one-cell cycle lag). Scale bar = 100 �m in F (applies in A?F). Inserts in (E,F) are amplified 9 � and 4.5�, respectively. Diagrams on the bottom left of each panel illustrate an idealized spindle during metaphase (microtubules in red, centrosome in green) within a blastomere at each stage. Spindles exhibit a normal bipolar structure in all panels except in the embryo undergoing a cell division stall in E, in which a microtubule-based monoaster emanates from a single centrosome. In the micrographs, the bipolar spindle exhibits somewhat different appearances due to its rapid reorganization during the cell cycle (Theusch et al., 2006; Yabe et al., 2007; W�hr et al., 2010): compact spindle midzone with weakly labeling asters during metaphase (as in C), solid asters during early anaphase (as in B, E, F), wider, hollow asters during late anaphase (as in A, D). Microtubule enrichments observed between parallel spindles in B, C, and F, or between monoasters in E, correspond to the furrow microtubule array for the previous cell cycle, which is derived from astral microtubules and forms during cytokinesis. |
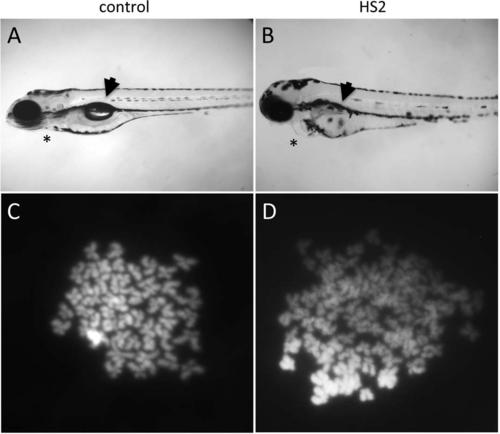
HS2 promotes whole genome duplication and tetraploidy. A,B: Control (A) and HS2-treated (B) embryos derived from in vitro fertilization with untreated sperm, at 5 dpf. In HS2-treated embryos, the embryonic cavity (asterisk) is distended compared with controls, and the swim bladder (arrowhead) is not inflated. C,D: Chromosome counts in cells from 24 hpf embryos. C: Normal diploid chromosomal content observed in cells from embryos exposed to normal temperatures and which exhibit no cell division stall. D: Tetraploid chromosomal content in cells from HS2-treated embryos and which showed a cell division stall in the second cell cycle. |
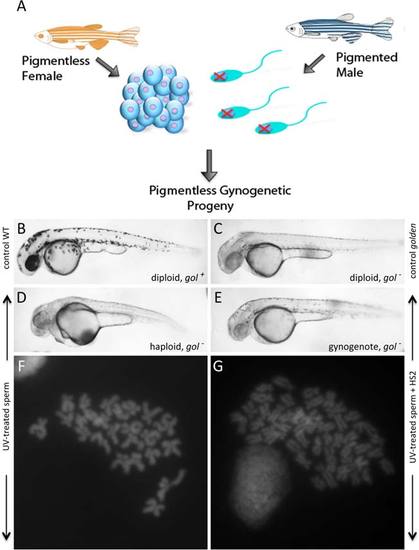
HS2 coupled to haploid production induces gynogenetic diploids. A: Diagram of the gynogenetic procedure. Maternal homozygosity for pigment markers is used to confirm that the resulting progeny lack genetic material from (wild-type pigmented) males. B?E: Phenotypes of 36 hpf embryos: (B) control wild-type (WT): diploid with normal (gol+) pigmentation, (C) control golden: wild-type diploid, homozygous for a mutation in the pigmentation gene golden (gol), which results in reduced levels of pigment in both melanophores and the eye, (D) haploid: haploid embryo derived from the fertilization of eggs from golden homozygous females with UV-treated sperm from wild-type males. The shortened body axis reflects the characteristic haploid phenotype. E: Gynogenote: gynogenetic diploid produced by treating haploid embryos (which would otherwise develop into embryos shown in C) with HS2 and selecting for a one-cycle division lag during the second cell cycle. In both D and E, golden pigmentation of the embryo indicates only maternal genetic material was transferred to the embryo. F,G: Chromosome counts in cells from 24 hpf embryos. F: Haploid chromosomal content in haploid embryos depicted in D. G: Diploid chromosomal content in gynogenetic diploids depicted in E. |
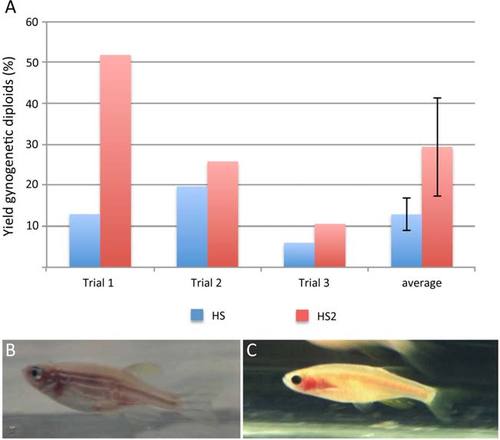
HS2 exhibits higher yields of gynogenetic diploids than standard Heat Shock. A: In three independent trials, pools of eggs were divided and treated with standard Heat Shock (HS) or HS2. HS2-treated embryos exhibit consistently higher yields of gynogenetic production. Bars indicate the standard error in average yield values. A chi-square test for equality of proportions performed on the combined data shows a significant difference in the average yield values between standard Heat Shock and HS2 (P-value = 0.016; ?2 = 5.8). Control untreated embryos resulted in 100% embryos with haploid morphology. B,C: HS2-derived gynogenetic adult offspring from golden (B) and casper (C) mutant lines. |