
- Title
-
A novel FoxD3 gene trap line reveals neural crest precursor movement and a role for FoxD3 in their specification
- Authors
- Hochgreb-H‰gele, T., and Bronner, M.E.
- Source
- Full text @ Dev. Biol.
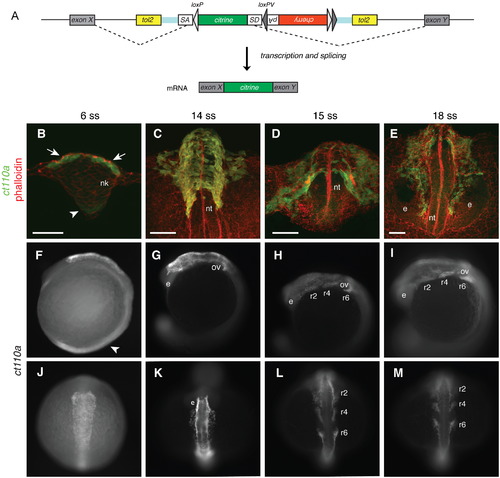
A novel protein trap line allows visualization of neural crest cells from early stages in zebrafish. (A) Schematic diagram of the FlipTrap strategy. The FlipTrap construct contains an internal exon, consisting of citrine sequence lacking start and stop codons (green), flanked by splice acceptor (SA) and donor (SD) sites. Genomic integration of the construct is mediated by Tol2 transposase. When the FlipTrap construct inserts within an intronic region, the endogenous splicing machinery recognizes SA and SD sites, and the citrine sequence is integrated into the mRNA (dashed lines), resulting in an endogenously expressed Citrine-fusion protein. (B?D) Confocal images of ct110a embryos. Transverse sections. C and D are 3D-projections of z-stacks. Embryos counterstained with phalloidin-rhodamine (red) and Citrine signal amplified by immunohistochemistry against GFP after fixation (green). (B) Expression of Citrine is detected in the dorso-lateral cells (white arrow) of the neural keel (nk) at 6-somite stage (ss). Low-level expression can be detected in the floor plate (white arrowhead). (C) Delamination of neural crest cells (NCCs) from the neural tube (nt). Citrine-positive cells distributed on the dorsal aspect of the neural tube at 14ss. (D) Migration of NCCs from the neural tube along stereotyped routes throughout the embryo. (E) At 18ss, cranial NCCs migrate anteriorly to the head, circumventing the eyes (e). Scale bars: 50 μm. (F?M) Expression of Citrine in ct110a embryos. (F?I) Left side view. (J?M) Dorsal view. Citrine signal amplified by GFP immunostaining in fixed embryos. (F and J) Expression of Citrine in NCCs on the dorsal aspect of the anterior neural tube, at the level of the hindbrain. Low expression in the floor plate region (white arrowhead). (G and K) NCCs delaminate and migrate from the neural tube to the head, around the otic vesicles (ov) and eyes (e). (H, I, L and M) NCCs migrate ventrally into the pharyngeal arches in three separate streams along stereotyped routes (r2, r4 and r6). EXPRESSION / LABELING:
|
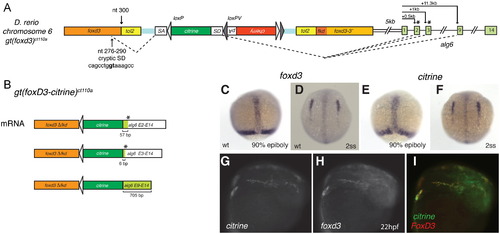
Expression of citrine in ct110a reflects endogenous expression of foxd3. (A) Schematic diagram of genomic FlipTrap integration in the ct110a line. Integration of the FlipTrap construct occurred within the coding region of foxd3, in chromosome 6. Splice acceptor site (SA) of the citrine internal exon recognizes a cryptic splice donor site within foxd3 sequence (nucleotides 276?290 of foxd3 coding sequence). The splice donor site (SD) of citrine reacts with acceptor sites of alg6. The citrine exon splices with exons 3 or 9 of alg6, respectively situated at +1 kb or +11.3 kb downstream from the alg6 start site in the genomic sequence. (B) Splicing events produce a chimeric Citrine-fusion protein, which encodes the N-terminal portion of FoxD3 (FoxD3??Fkd) and an Alg6 tail. Splicing of citrine with exon 2 or 3 of alg6 results in a short Alg6 tail (19 or 2 aminoacids, respectively), caused by a premature stop signal (*) generated by a frame shift. Alternative splicing with exon 9 does not affect the coding frame. (C?I) Expression of citrine recapitulates foxd3 expression in the Gt(foxd3-citrine)ct110a line. Whole mount in situ hybridization to foxd3 (C and D) and citrine (E and F). (C) Early expression of foxd3 in the prechordal plate and blastoderm margin of zebrafish embryos in late gastrula. (D) foxd3 expression by neural crest cells initiates during early somitogenesis, as bilateral fields in the anterior neural plate border. (E and F) Distribution of citrine transcripts in Gt(foxd3-citrine)ct110a embryos corresponds to endogenous expression of foxd3. (G?I) citrine mRNA co-localizes with foxd3 endogenous expression in neural crest cells. Double fluorescent whole mount in situ hybridization to citrine and foxd3 in Gt(foxd3-citrine)ct110a embryos at 22 hpf. EXPRESSION / LABELING:
|
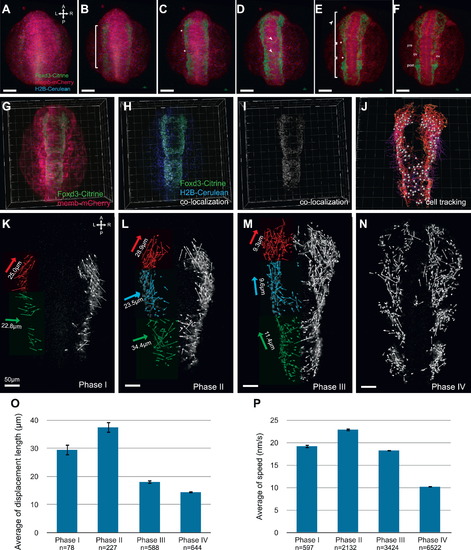
Analysis of the dynamics of neural crest migration in ct110a embryos. (A?F) Confocal projections of time-lapse imaging of live Gt(foxd3-citrine)ct110a embryos. Dorsal views. Panels represent 1 h intervals. FoxD3-Citrine signal is depicted in green; membrane-tagged mCherry, red; H2B-Cerulean, blue. (A) Onset of FoxD3-Citrine expression at the 1?2 somite stage as NCCs are induced in the anterior neural plate border. (B) Expansion of FoxD3-Citrine expression along the neural plate border. (C) Segmentation of the neural crest field is first delineated by the appearance of furrows (asterisks) along its rostral-caudal extent. (D) A subset of NC cells moves towards the midline (arrowheads) at specific anterior-posterior levels. (E) Citrine-positive cells return from the midline to the lateral aspect of the neural tube, migrate towards the head and branchial arches, where they will condense. (F) Consolidation of the rostral-caudal segmentation of cranial neural crest as they migrate anteriorly to the head region, or condense in pre- and post-otic ganglia. Scale bars, 100 μm. (G?J) Cell tracking analysis of time-lapse dataset. Dorsal views. (G) Panel represents projection of a single time-point. (H and I) Co-localization of Citrine signal with H2B-Cerulean is depicted in white. (J) Tracing of NC migration obtained by tracking co-localization signal in a representative time-lapse dataset of Gt(foxd3-citrine)ct110a embryos. (K?P) Phases of the morphogenetic movements of cranial NCCs in zebrafish. (K?N) Projection of NC displacement trajectories analyzed for each phase. Dorsal views. Thin arrows represent the displacement vectors of NC cells. Thick arrows are the projection of the resultant displacement unit vector, color-coded for each corresponding region. Resultant magnitude values are normalized for the number of trajectories and indicated in micrometers. Scale bars 50 μm. (K) Induction of NCCs in the neural plate border and initial migration to the midline. (L) Migration of NC to the midline and anteriorly. (M) NCCs migrate laterally and anteriorly towards the branchial arches and head. (N) Consolidation of NCCs position in branchial arches and head region for subsequent differentiation. (O) Quantitative analysis (mean±s.e.m.) showed that on average, displacement of cranial NC cells is significantly distinct during Phases I, II and III/IV. Difference in displacement length between phases III and IV is not significant. ANOVA statistical test P<0.0001. (P) Quantitative analysis showed that NC migrate faster during Phase II, and slow down as cells migrate to their final destination for subsequent differentiation. Speed is significantly different among Phases I through IV. ANOVA test P<0.0001. EXPRESSION / LABELING:
|
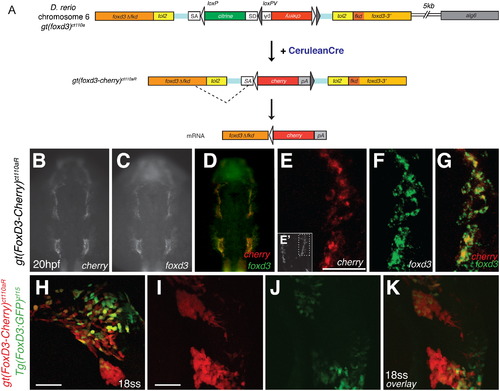
Cre-mediated recombination of Gt(foxd3-citrine)ct110a results in expression of a mutant FoxD3-mCherry fusion protein. (A) Schematic diagram of Cre-loxP recombination in Gt(foxd3-citrine)ct110a zebrafish line. The FlipTrap construct contains inverted mCherry (red box) and polyA (pA) sequences, flanked by heterotypic loxP (light gray) and loxPV (dark gray) sites. In embryos injected with cerulean-cre mRNA, recombination of the loxP sites results in deletion of citrine and correct orientation of mcherry-pA. Splicing of foxd3 cryptic splice donor site with mcherry splice acceptor site (SA) produces a mutant FoxD3?Fkd-mCherry fusion protein, expressed under foxd3 endogenous regulation. (B?K) Expression of mcherry co-localizes with endogenous foxd3 in Gt(foxd3-mcherry)ct110R embryos, whereas Tg(foxd3:GFP) line recognizes a subpopulation of NCCs. (B?G) Fluorescent whole mount in situ hybridization in Gt(foxd3-mcherry)ct110aR embryos at 22 hpf. Expression of mcherry (B) is detected in the branchial arches, in the same distribution as endogenous foxd3 (C and D). High-resolution confocal imaging of mcherry (red; E and G) and foxd3 (green; F and G) shows co-expression in the branchial arches, as shown in the inset (E′, dashed line). (H?K) Confocal imaging of double transgenic Gt(foxd3-mcherry)ct110aR; Tg(foxd3:GFP) embryo at 18-somite stage. Expression of ct110aR (red) allows visualization of early NCCs associated with the neural tube (K) and NC in the branchial arches (H). In contrast to expression of FoxD3-mCherry, which comprehensively labels neural crest cells, expression of GFP (green) driven by a 14 kb promoter is heterogeneously distributed (H and J) and shows restricted expression in the branchial arches at 18ss (J and K). Scale bars: 50 μm. EXPRESSION / LABELING:
|
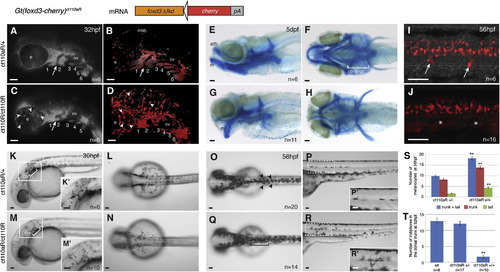
FoxD3 loss-of-function in Gt(foxd3-mcherry)ct110aR homozygotes reveals multiple aspects of FoxD3 mutant phenotype. Development of Gt(foxd3-mcherry)ct110aR heterozygotes (ct110aR/+) is normal. Homozygosis (ct110aR/ct110aR) results in a FoxD3 mutant phenotype. (A?D) Loss of FoxD3 affects migration of NCCs to the pharyngeal arches. Six embryos with each genotype were live imaged and analyzed, showing similar phenotypes. Lateral views, anterior to the left. (A and C) Images of mCherry fluorescence in the head of Gt(foxd3-mcherry)ct110aR embryo. (B and D) Confocal projections of Gt(foxd3m-mcherry)ct110aR embryos head region. Expression of FoxD3-mCherry allows visualization of NCCs in the branchial arches of Gt(foxd3-mcherry)ct110aR embryos. (A) Normal development and segmentation of pharyngeal arches (1?6) in Gt(foxd3-mcherry)ct110aR heterozygotes. (B) Confocal imaging of Gt(foxd3-mcherry)ct110aR heterozygous embryo shows FoxD3-positive cells sparsely distributed in a ring-like arrangement around and on top of the eyes (e), and outlining the midbrain-hindbrain boundary (mhb). (C) In Gt(foxd3-mcherry)ct110aR homozygotes, segmentation between the first (1) and second (2) pharyngeal arches is disrupted (white arrow). Posterior arches (3?6) are present, but show reduced sizes in homozygotes. (D) Confocal imaging shows an overall increase in mCherry-positive cells in Gt(foxd3-mcherry)ct110aR homozygotes. Note mesenchymal-like aspect of mCherry-positive cells, with less adhesion and more cytoplasmic protrusions. In particular, large cells are widely distributed throughout the eyes and brain (arrowhead). (E?H) Morphogenesis of the jaw cartilage is disrupted in foxd3 mutants. Alcian Blue staining of Gt(foxd3-mcherry)ct110aR embryos at 5 dpf. (E,G) Lateral view, anterior to the left. (F,H) Ventral views. (E and F) Craniofacial cartilage develops normally in Gt(foxd3-mcherry)ct110aR homozygotes. (G) Cartilage defects in Gt(foxd3m-mcherry)ct110aR homozygotes are characterized by ventral projection of the Meckel′s cartilage (m) and ethmoid plate (eth). Derivatives of the second arch (ceratohyal, ch and hyosymplectic, h), as well as ceratobranchials I?V (cb i?v), are hypoplastic and malformed. (I and J) Formation of DRGs is disrupted in Gt(foxd3-mcherry)ct110a homozygous embryos. (I and K) Confocal projection of mCherry-positive cells in the trunk of live Gt(foxd3-mcherry)ct110a embryos at 56 hpf. DRGs emerge from the neural tube at the level of the anterior half of each somite. (I) FoxD3 is expressed in the glial cells associated with the DRGs (white arrows). (J) Gt(foxd3-mcherry)ct110a homozygotes consistently lack DRGs and associated glia (white asterisks), disrupting the periodicity of DRG emergence from the neural tube. (K?N) Specification of melanophores is delayed in FoxD3 mutants. Live embryos at 30 hpf, anterior to the left. (K and M) Lateral views. (L and N) Dorsal views. (K and L) Melanophores concentrate posterior to the otic vesicle (ov), from where they migrate over the yolk. (M and N) Melanophores are consistenly reduced in Gt(foxd3-mcherry)ct110a homozygotes at 30 hpf. (K′,M′) Higher magnification of the boxed area. (O?S) FoxD3 is a repressor of the melanocytic lineage at the expense of iridophore cells. (O and Q) Dorsal view of live embryos, anterior to the left. (P and R) Left lateral view, trunk region. (O) Melanophores are distributed in a characteristic pattern in the zebrafish embryo at 56hpf. They can be individually visualized along the dorsal part of the trunk and tail (black arrowheads). (P and P′) Melanophores are also distributed as flat cells along the lateral line, mostly at the level of the trunk. (Q) In Gt(foxd3-mcherry)ct110a homozygotes, dorsal melanophores are increased in the trunk, but not in the head region. (R) Melanophores in the lateral line are increased in homozygotes (n=14), with greater incidence of cells in the tail region (black asterisks). Melanophores appear dislocated from the lateral line and present increased cytoplasmic protrusions (R′). (S) Quantification of melanophores in the trunk and tail region of embryos at 56 hpf. Significant increase was observed in homozygotes. **P<0.001. (T) Quantification of ridophores in the dorsal trunk of embryos at 52 hpf. Iridophores are significantly reduced in homozygous Gt(foxd3-mcherry)ct110a at 52 hpf. **P<0.0001. Scale bars: 50 μm. PHENOTYPE:
|
Reprinted from Developmental Biology, 374(1), Hochgreb-H‰gele, T., and Bronner, M.E., A novel FoxD3 gene trap line reveals neural crest precursor movement and a role for FoxD3 in their specification, 1-11, Copyright (2013) with permission from Elsevier. Full text @ Dev. Biol.