
- Title
-
The ceramide synthase 2b gene mediates genomic sensing and regulation of sphingosine Levels during zebrafish embryogenesis
- Authors
- Mendelson, K., Pandey, S., Hisano, Y., Carellini, F., Das, B., Hla, T., Evans, T.
- Source
- Full text @ Elife
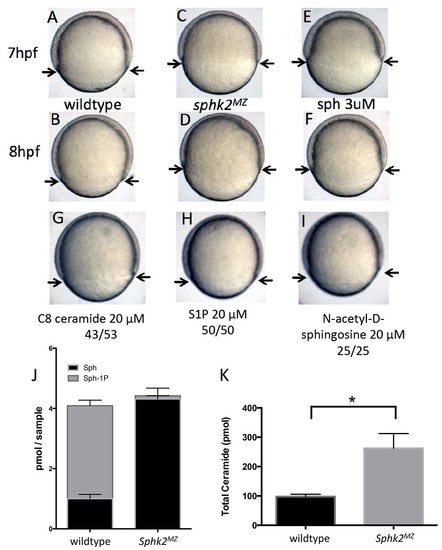
The sphk2MZ embryos show developmental delay associated with enhanced levels of sphingosine. (A,B) Compared to time-matched representative wildtype embryos observed at 7 or 8 hr post fertilization (hpf), the sphk2MZ embryos (C,D) display a developmental delay during epiboly, denoted by arrows indicating the leading edge of the blastoderm. (E, F) Wildtype embryos treated at 2 hpf with 3 ?M sphingosine (sph) were similarly developmentally delayed. Treatment of wildtype embryos with G) C8 ceramide (20 ?M), (H) S1P (20 ?M), or (I) N-acetyl-D-sphingosine (20 ?M) had no adverse effects on epiboly. A-I show representative embryos, n at least 25 in each of 3 independent experiments. (J) Based on lipidomic profiling at 6 hpf, the sphk2MZ embryos are depleted of S1P (Sph-1P) and have elevated levels of sphingosine and (K) total long-chain ceramides compared to time-matched wildtype embryos. For J and K, results are the average of three independent experiments pooling 50 embryos each; errors bars indicate the standard error of the mean. PHENOTYPE:
|
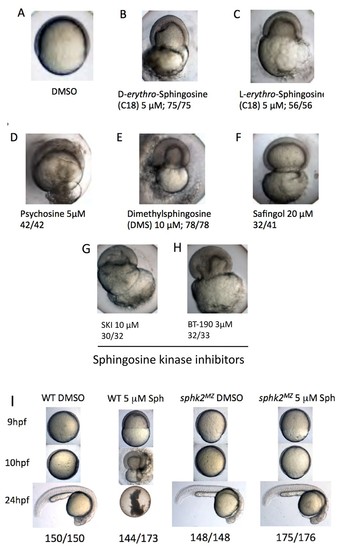
Excess sphingosine results in gastrulation defects and embryonic lethality, which is abrogated in the context of a sphk2MZ mutation. (A) Embryos cultured in DMSO display normal coverage of the yolk by the developing blastoderm during epiboly at 7 hpf. (B) Treatment of wildtype embryos with 5 ?M D-erythro-sphingosine (the active naturally occurring isomer) or (C) 5 ?M L-erythro-sphingosine (the inactive isomer) caused stalling of embryonic development during epiboly, rupture of the yolk membrane, and failure of the embryos to complete gastrulation. This phenotype was recapitulated by embryos treated with (D) 5 ?M D-galactosyl-�1?1'-D-erythro-sphingosine (Psychosine), (E) 10 ?M dimethylsphingosine (DMS), and (F) 20 ?M L-threo-Dihydrosphingosine (Safingol). Treatment of wildtype embryos with two chemical inhibitors of sphingosine kinases, (G) SKI (10 ?M) or (H) BT-190 (3 ?M) recapitulated the catastrophic embryonic phenotype. (I) The majority (144/173, 83%) of wildtype (WT) embryos treated with 5 ?M sphingosine exhibited a lethal phenotype during gastrulation, whereas the majority (175/176, 99%) of the sphk2MZ embryos treated with 5 ?M sphingosine completed gastrulaton and early development, only subsequently displaying the cardiac bifid phenotype observed in sphk2MZ embryos, with no additional defects. Control embryos were cultured in DMSO vehicle (1%). Shown are representative embryos, n is indicated, from at least three independent experiments. PHENOTYPE:
|
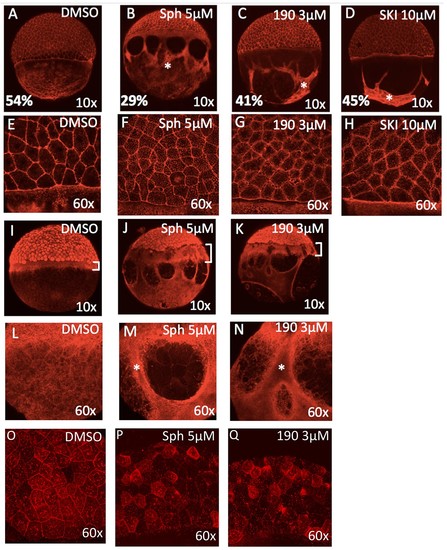
Excess sphingosine disrupts actin and microtubule organization and reduces junctional E-cadherin. (A) During gastrulation, actin accumulates at the vegetal pole as shown by phalloidin staining in DMSO-treated control embryos at ~50% epiboly (5.3 hpf). Phalloidin staining shows aggregation of thick F-actin positive bundles within the yolk cytoplasmic layer (YCL) of embryos treated with (B) 5 ?M sphingosine, (C) 3 ?M BT-190, or (D) 10 ?M SKI (indicated by asterisks). For A-D, the average percent progression of the EVL is indicated as measured from the pole for a representative set of 5 embryos for each condition. (E?H) Shown at higher magnification, F-actin also accumulates within the cytoplasm of the enveloping cells in the treated embryos compared to DMSO-treated controls. During epiboly, two perpendicular microtubule layers are visible in DMSO-treated embryos at the YSL (I) and in the YCL (L). Staining with ?-tubulin revealed a massively thickened microtubule layer within the YSL (as indicated by brackets, which also highlights the delayed progression of the EVL compared to deep layer cells) in embryos treated with (J) 5 ?M sphingosine or (K) 3 ?M BT-190 as well as (M,N) microtubule aggregation in certain areas of the YCL (denoted by asterisks) and other areas that are completely devoid of microtubules. Indicated by brackets in (I-K), the progression of the EVL appears to be more delayed compared to the deep cell layer. (O) DMSO-treated embryos display junctional E-cadherin staining of the EVL cells. (P,Q) Reduced junctional E-cadherin staining of the EVL cells and accumulation of staining within the EVL cytoplasm in embryos treated with 5 ?M sphingosine or 3 ?M BT-190. For all panels, representative embryos are shown, n is at least 100 from at least three independent experiments. PHENOTYPE:
|
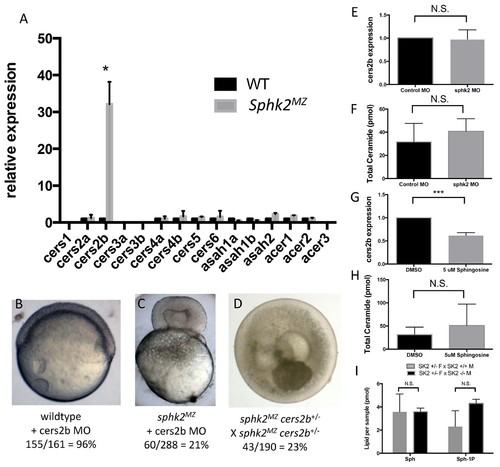
The sphk2MZ embryos express strikingly high levels of cers2b transcripts. (A) Analysis of ceramide synthase and ceramidase genes by qRT-PCR shows that cers2b is significantly upregulated in sphk2MZ embryos (p=0.019) compared to wildtype embryos at 2 hpf. (B) Representative wildtype or (C) sphk2MZ embryos following injection with 5.3 ng of a cers2b translation blocking morpholino (MO), alongside (D) embryos derived from a sphk2MZ; cers2b+/- incross. For wildtype embryos, 155/161 (96%) completed normal gastrulation, epiboly, and appeared normal at 2 dpf. Following injection into sphk2MZ embryos, 60/288 or 21% from 4 separate experiments recapitulated the early gastrulation lethality phenotype (p=0.0154). Correspondingly, 43/190 or 23% of embryos derived from crossing sphk2MZ; cers2b+/- adult fish recapitulated early gastrulation lethality phenotype by 9 hpf. (E) Sphk2 morphant embryos fail to upregulate cers2b transcripts and (F) did not vary in ceramide levels when compared to those injected with control morpholino. (G) Embryos exposed to 5 uM sphingosine in culture similarly fail to upregulate cers2b transcript levels and (H) showed no differences in ceramide accumulation. (I) There were no significant differences in sphingosine (Sph) or S1P (Sph-1P) levels in embryos with maternally deposited Sphk2, consistent with normal development. Embryos were harvested for both qPCR and lipid measurements at 6 hpf. All experiments were repeated at least three times with equivalent results. EXPRESSION / LABELING:
PHENOTYPE:
|
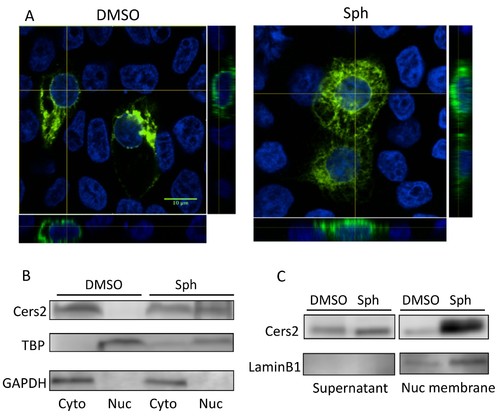
The subcellular localization of Cers2 protein is altered by increased sphingosine levels. HEK293T cells transfected with a Cers2-GFP expression plasmid were examined after 6 hr exposure to 10 uM sphingosine in culture. (A) Cells treated with sphingosine (Sph) show subcellular relocalization of Cers2b-GFP (green) with more overlap within nuclear regions (DAPI, blue) as seen in the z-stack (side panels). (B?C) Western blotting analysis of lysates probed with antibodies specific to Cers2, GAPDH (as a control for cytoplasmic proteins), TBP (as a control for nuclear proteins) or LaminB1 (as a control for nuclear membrane associated proteins). (B) Subcellular fractionation into cytoplasmic (Cyto) and nuclear (Nuc) lysates confirms increased nuclear association of Cers2 in cells treated with sphingosine. (C) Fractionation using a nuclear membrane isolation kit indicates Cers2 is most enriched in the nuclear membrane fraction. |
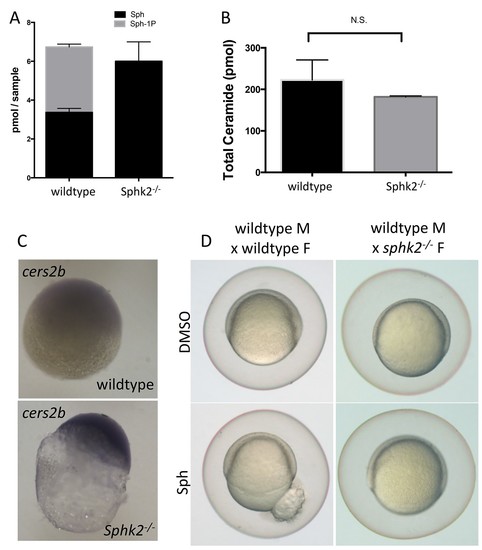
Transcriptional changes during oogenesis underlie sphk2MZ resistance to sphingosine buildup. Oocytes isolated from wildtype and sphk2-/- female fish show (A) increased sphingosine (Sph) levels in mutant oocytes associated with loss of S1P (Sph-1P) with (B) no difference in total ceramide levels. (C) Oocytes derived from sphk2-/- oocytes showed significant upregulation of cers2b transcripts, as imaged following in situ hybridization. Note that signal in the wildtype oocytes was equivalent to background staining using a sense-strand control probe. (D) Correspondingly, embryos derived from sphk2-/- females (right) showed relative tolerance to sphingosine accumulation when compared to those from wildtype females (left) when cultured in medium with 5 uM sphingosine. For (A, B) . results are averaged from three independent experiments using at least 50 embryos per measurement. For (C,D), representative embryos are shown, n > 25, from at least three independent experiments. EXPRESSION / LABELING:
PHENOTYPE:
|