
- Title
-
Bmp inhibition is necessary for post-gastrulation patterning and morphogenesis of the zebrafish tailbud
- Authors
- Row, R., and Kimelman, D.
- Source
- Full text @ Dev. Biol.
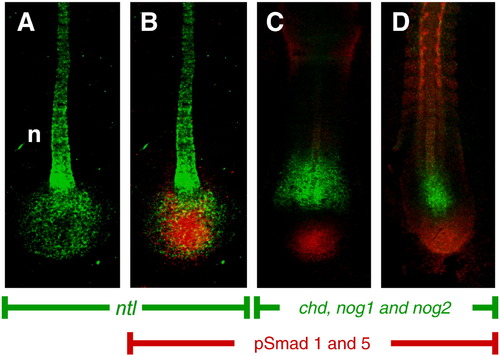
Bmp signaling is tightly regulated in the tailbud during early somitogenesis. (A, B) Bmp signaling is active in the mesodermal progenitor zone. Expression of no tail is shown by FISH in green in 3-somite stage embryos marking both the mesodermal progenitor zone and the notochord (n). Bmp signaling is indicated by antibody staining for phosphorylated Smad proteins (red in B?D). Bmp inhibitors are expressed just anterior to the activated Smad zone. The Bmp inhibitors chordin, noggin1 and noggin2 are shown by FISH in green and the Smad staining is shown in red in 3-somite (C) and 12-somite (D) embryos. All embryos are shown from a dorsal view with anterior to the top. EXPRESSION / LABELING:
|

A new transgenic line allows temporal cell-autonomous activation of Bmp signaling. (A) Schematic diagram of construct used to create the new line. A multimerized minimal heat shock-responsive promoter is flanked by genes for GFP and a constitutively active mutant form of the Type Ib Bmp receptor. Tol2 elements enhance transgenesis efficiency. (B) Early heat shock of a transgenic embryo at dome stage recapitulates the expected Bmp ventralized phenotype. (C) eve1 expression is rapidly induced by ectopic Bmp signaling. Embryos were heat-shocked at dome stage and fixed at shield stage. |

Ectopic Bmp signaling during early somitogenesis alters tailbud morphogenesis. (A?D) Transgenic embryos (right) and wild-type controls were heat shocked at the 3-somite stage. No morphological change is visible at the 9-somite stage (B). By the 16-somite stage the tailbud and posterior somites display clear alterations (D). (E, F) At approximately 42 hpf transgenic embryos (F) display a shortened body axis and the epidermal fin fold has failed to form normally. PHENOTYPE:
|
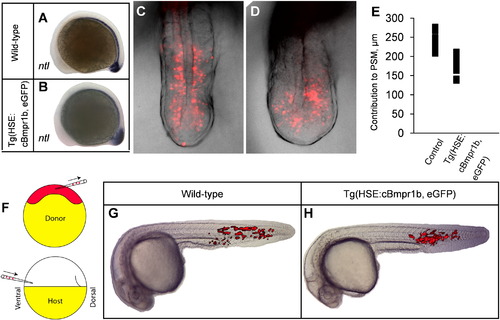
Ectopic Bmp signaling during early somitogenesis alters patterning and morphogenesis. (A, B) Expression of no tail is expanded in the tailbud of transgenic embryos at the 9-somite stage. (C?E) Cell migration, as shown by photoconverted NLS-Kikume fluorescence (red), is disrupted as a result of ectopic Bmp signaling. The graph shows the most anteriorward migration of labeled cells from the posterior end of the embryo in individual embryos. (F, G, H) Cell migration defects are a non-cell-autonomous result of ectopic Bmp signaling. Cell transplants were performed as diagrammed in (F). Cells from a transgenic donor (H) contribute to the same cell types and anterior extent as those from a wild-type donor (G). Embryos were heat shocked at the 3-somite stage. EXPRESSION / LABELING:
PHENOTYPE:
|
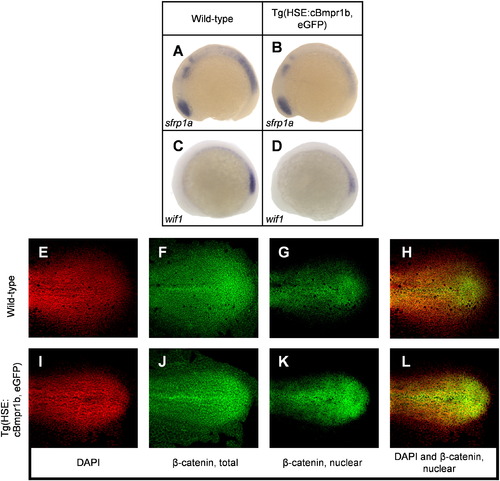
Ectopic Bmp signaling enhances Wnt signaling. (A?D) Ectopic Bmp signaling represses expression of secreted Wnt inhibitors sfrp1a and wif1. (E?L) Ectopic Bmp signaling expands the region of canonical Wnt signaling in the tailbud. Nuclear β-catenin accumulates in a larger region in transgenic embryos after heat shock. EXPRESSION / LABELING:
|
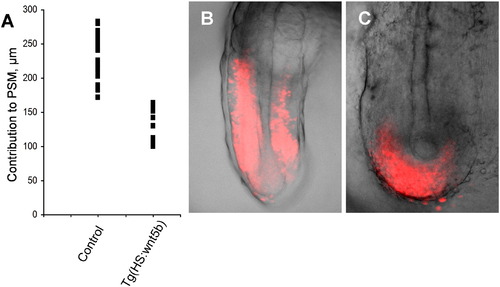
Ectopic Wnt5b expression recapitulates ectopic Bmp signaling in the tailbud. Cell migration, as shown by photoconverted NLS-Kikume fluorescence (red), is disrupted as a result of ectopic Wnt5b expression. The graph shows the most anterior contribution of labeled cells to presomitic mesoderm in individual embryos. |
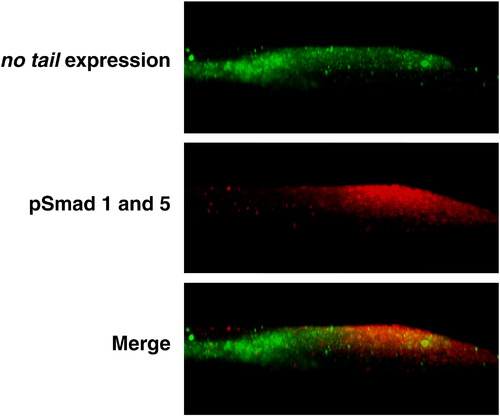
Bmp signaling is active in the mesoderm progenitor zone. Active Bmp signaling (red) is principally active in the dorsal-posterior region of the tailbud, where expression of no tail (green) marks the mesoderm progenitor zone. The strongest ntl expression is in the notochord (see Fig. 1A). Shown is a confocal section at the midline of a 3-somite tailbud with dorsal at top and posterior at right. |

Ectopic Bmp signaling disrupts somite morphogenesis. Wild-type (top) and transgenic (bottom) embryos were heat shocked at the 3-somite stage and fixed at approximately 32 h post fertilization. DAPI staining shows the nuclei aligned along clear borders in the chevron-shaped somites of the wild type embryo. Anterior somites in the transgenic embryo have indistinct or nonexistent borders. Posterior somites show partially normal segmentation but do not form the correct shape. |
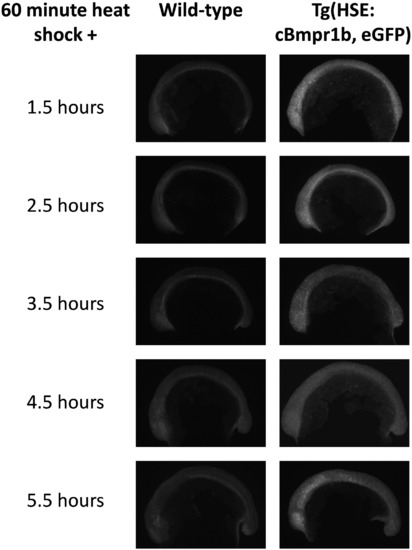
Ectopic Bmp signaling is transiently induced by heat shock. Wild-type (left) and transgenic (right) embryos were heat shocked at the 3-somite stage and fixed at the indicated intervals. All were stained for the presence of phosphorylated Smads 1 and 5. In wild type embryos p-Smad is restricted to the tailbud and developing somites. Transgenic embryos show high levels of p-Smad throughout all tissues within 90 min of heat shock activation. This appears to peak approximately 2.5 h after the heat shock ends. |

Ectopic Bmp signaling minimally affects expression of bmp2a and bmp4. Wild-type (left) and transgenic (right) embryos were heat shocked at the 3-somite stage and fixed at the 9-somite stage, then stained for expression of bmp2a or bmp4 by WISH. No change in the expression of bmp2a is observed. The region of bmp4 expression in the tailbud is enlarged very slightly, and a small number of epidermal cells express this ligand ectopically. |

Inhibitors of Bmp and Wnt signaling respond to ectopic Bmp signal. Transgenic and control embryos were heat shocked at 3 somites and fixed at 9 somites. In-situ hybridization demonstrates that ectopic Bmp signaling upregulates expression of noggin1 and noggin2. Expression of chordin may be very slightly downregulated. wif1 expression is reduced but not eliminated. It is shown in combination with pax2.1 staining, which does not change under these conditions. |
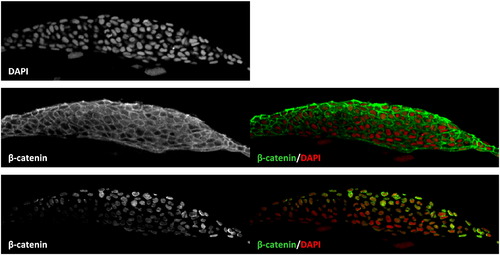
Nuclear β-catenin can be measured by comparison to DAPI staining. Optical section of a tailbud imaged laterally, stained with DAPI and antibody against β-catenin. The high levels of β-catenin at the cell membrane obscure other staining within the cell. Using the computer program ImageJ to define the area of the nuclei based on DAPI staining allows for specific exclusion of non-nuclear staining. The resulting composite clearly shows regions of active canonical Wnt signaling. |
Reprinted from Developmental Biology, 329(1), Row, R., and Kimelman, D., Bmp inhibition is necessary for post-gastrulation patterning and morphogenesis of the zebrafish tailbud, 55-63, Copyright (2009) with permission from Elsevier. Full text @ Dev. Biol.